Plant Breeding: Germplasm Resources
A. Genetic Base and Diversity
Expeditions and collections made by early oil palm researchers at the various centers in West Africa e.g Nigeria (Calabar, Ufuma), Ivory Coast (La Me), Zaire (Sibiti, Yangambi,), especially after World War II formed the genetic base and diversity of the respective oil palm breeding programs (Rosenquist 1986; Hartley 1988). There were subsequent genetic material exchanges for breeding purposes. Most of the oil palm breeding populations were descended from one or a few palms and were termed by Rosenquist (1986) as breeding populations of restricted origins. A brief description of some of the major breeding populations is given below:
1. Deli. This is the thick-shelled dura variety derived from the original four Bogor palms in Java. Distribution of the subsequent progenies to other countries followed by local selection led to the development of the Elmina, Serdang Avenue, and Ulu Remis Deli dura subpopulations/selections in Malaysia and the Dabou and La Me dura subpopulations/selections in Ivory Coast. The Ulu Remis Deli duras were most widely distributed. The rather uniform high yielding Deli population led to the speculation of a common progenitor for the four Bogor palms. Deli duras provide the mother palms for almost all major oil palm commercial hybrid seed production programs. The Dumpy and Gunung Melayu palms are short variants of the Deli;
2. AVROS . Seeds from the Djongo (best) palm at Eala Botanical Garden in Zaire were obtained and planted in 1923 by Algemeene Vereniging van Rubberplanters ter Oostkust van Sumatra (AVROS) at Sungai Pancur to give rise to the well-known SP540 tenera palm. Crossing with the teneras at Bangun Bandar Experimental Station and subsequent backcrossing to the SP540 selfs resulted in the AVROS population. AVROS pisiferas are noted for their vigorous growth, precocious bearing, thin shell, thick mesocarp and high oil yield conferring attributes. Major commercial hybrid seed production programs in Colombia, Costa Rica, Indonesia, Malaysia, and Papua New Guinea are based on Deli dura x AVROS pisifera lineage;
3. Yangambi . The Institut National pour l’Etude Agronomique du Congo (INEAC) started the breeding program at Yanagambi, Zaire with open-pollinated seeds from the Djongo palm and from teneras in Yawenda, N’gazi and Isangi. Their breeding led to the development of the Yangambi population characterized by its excessive vigor, bigger fruit and high oil yield conferring attributes. The Yangambi population is featured in many breeding and seed production programs worldwide;
4. La Me . The Institut de Recherches pour les Huiles et Oleagineux (IRHO) developed the La Me population from 21 tenera palms, particularly the Bret 10 palm, grown from seeds collected from the wild groves in Ivory Coast. Pisiferas and teneras derived from the L2T tenera palm are used in breeding and seed production programs in West Africa and Indonesia. La Me teneras and their progenies are characteristically smaller palms with smaller bunches and fruits but they appear to be more tolerant of sub-optimal growing conditions.
5. Binga . This subpopulation is derived essentially from the F 2 and F 3 Yangambi progenies planted in Binga Plantation, Yangambi, Zaire. Palms Ybi 69MAB and Bg 312/3 are the parent palms of breeding interest.
6. Ekona . The Ekona population is derived from the wild palms in the Ekona area of Cameroon and bred further in the Unilever plantations of Cowan Estate, Ndian Estate, and Lobe Estate. Progenies from palm CAM. 2/2311, noted for its high bunch yield, good oil content and wilt resistance, have been distributed to Costa Rica and Malaysia;
7. Calabar . The breeding populations of the Nigerian Institute for Oil Palm Research (NIFOR) are much broader based with collections from Aba, Calabar, Ufuma, Umuabi. Progenies from the Calabar selections were most interesting and have been distributed to Costa Rica, Ghana, Indonesia, Ivory Coast and Malaysia. Palm NF 32.3005 was featured in most of the distributions.
Despite the reasonably diverse original genetic base, the bulk of the current commercial oil palm hybrid seed production and the breeding programs that support them are still based on the Deli and the Yangambi/AVROS and La Me breeding populations which can be traced to one or two palms from the natural groves. Because of the high selection intensities practiced and the need to retain the proven pedigrees of the hybrids for commercial reasons, genetic variability has been rapidly diminished as stressed by a number of researchers (Thomas et al. 1969; Ooi et al. 1973; Hardon et al. 1987). Prompted by this and the increasing importance of the crop, Malaysia with the cooperation of Nigeria under the auspices of the International Board For Plant Genetic Resources made the first systematic large-scale collections in the semi-wild groves in the oil palm belt of Nigeria (Rajanaidu et al. 1979; Obasola et al. 1983) with the objectives of genetic conservation and base broadening. The study on the genetic structure of the natural populations from the Nigerian collection revealed greater variability within families and between families than between populations. This guided the approach made in subsequent collections in Angola, Cameroun, Gambia, Guinea, Madagascar, Sierra Leone, Tanzania, and Zaire (Rajanaidu and Rao 1988). There was also a limited attempt to capture different ecotypes. The collected materials were planted and maintained as living collections in Malaysia with a sample retained by the host country for each collection (Rajanaidu 1990b; Rajanaidu and Jalani 1994) although long term conservation through cryopreservation and in vitro methods has also been attempted (Engelmann and Duval 1986; Paranjothy et al. 1986; Rohani and Paranjothy 1995).
Collections have also been made for E. oleifera in South America (Brazil, Colombia, Costa Rica, Honduras, Panama, and Surinam as well as for other oil bearing palms e.g. Bactris gasipaes (Pejibaye), Jessenia-Oenocarpus,, Orbignya martiana (Babassu), which have unusual fatty acid composition and other uses. These could be used in future genetic modification of palm oil.
B. Base Broadening
Private breeding programs although obliged to broaden the genetic base of their breeding populations to ensure breeding progress are generally reluctant to outcross with less advanced breeding materials and thus tend to end up with related materials. High unsaturated fatty acid content, large kernel, and dwarf high yielding lines have been developed from the Nigerian collection and distributed to the private industry breeders. Private breeders are reluctant to exploit them directly in commercial hybrid seed production although they have been urged to do so (Rajanaidu et al. 1999, 2000), but prefer to introgress the desirable features into their advanced breeding parents to maintain the integrity, uniformity, and consistency of their commercial hybrids (Sharma 1999; Soh et al 1999). These qualities may not be assured in the commercial hybrids developed directly from the early selections of the semi-wild Nigerian collections.
C. Genetics and Cytogenetics
Most oil palm breeding programs being commercially oriented cannot afford to undertake basic genetic studies. Thus such studies tend to be incidental, derived from breeding trials and seldom meet the basic assumptions required for genetic analyses.
1. Qualitative Traits . The revelation of the simple Mendelian inheritance of the shell thickness trait in the palm fruit in the 1940s by Bernaert and Vanderweyen (1941) provided the basis of modern oil palm breeding and hybrid seed production schemes (Plate 1A, Figs. 2, 3). The dura palm is homozygous dominant for the thick shell trait while the pisifera palm is the shell-less recessive homozygote which is usually female sterile as its pistillate inflorescence tends to abort. The dura × pisifera hybrid is the thin-shelled tenera heterozygote and is the commercial variety. The duras which are the female parents in hybrid seed production are usually regenerated (100%) from dura × dura crosses while the pisifera male parents are regenerated from tenera × tenera (25% pisifera segregants) and tenera × pisifera (50% pisifera segregants) crosses. Dura × tenera crosses used in progeny-tests segregate 1:1 for dura and tenera palms. Fertile pisiferas do occur occasionally and fertile pisifera × fertile pisifera crosses produce 100% pisifera progenies. Although the presence of shell is under monogenic control, the thickness of the shell was purported to be modified by minor genes (Van der Vossen 1974). Okwuagwu (1988) and Okwuagwu and Okolo (1992,1994) postulated maternal inheritance of kernel size with the involvement of a kernel-inhibiting factor.
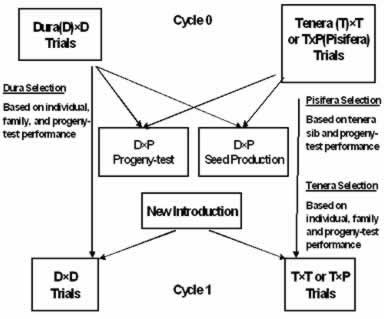
Fig. 2. Modified recurrent selection scheme.

Fig. 3. Modified reciprocal recurrent selection scheme. To sustain genetic variability for longer term improvement a recombinant phase involving outbred parents is run in parallel in a similar manner.
A number of other qualitative traits have also been reported from observations and genetic analyses to be simply inherited (Hartley 1988) e.g. crown disease, leaf form (idolatrica = fused pinnae), fruit shape (mantled), and fruit color (nigrescens = black unripe, virescens = green unripe, albescence = whitish unripe). Although the Dumpy trait appeared to be simply inherited, observations on segregating progenies did not confirm it (Soh et al. 1981). The mode of inheritance for tolerance to fusarium wilt has yet to be elucidated (De Franqueville and Renard 1990).
2. Quantitative Traits . Estimations of genetic variance and heritability have been attempted by oil palm breeders to provide information to guide them to increased breeding efficiency. This involves information on the existence of sufficient genetic variability, the choice of the population to improve and the adoption of appropriate breeding procedure to obtain the best selection response (Soh and Tan 1983). North Carolina Model 1 (hierarchical) and Model 2 (factorial) mating designs and the parent-offspring method are commonly used to estimate genetic variance and heritability in oil palm. Heritability estimates for various agronomic traits have been compiled by Soh and Tan (1983) and Hardon et al. (1985). With the use of data from breeding experiments derived from selected parents from very restricted populations, some of the assumptions such as random choice of parents, no correlation of genotypes at separate loci, and a definite reference population, necessary for valid quantitative genetic analyses could not be met and the biases incurred could not be estimated. These estimates merely serve as a guide for selection decisions as any estimate of heritability is unique i.e. dependent on the population and environment sampled and the estimation method used. For selection decisions or for prediction of selection response for a particular population and environment, specific heritability estimate obtained from the population concerned should be used. In lieu of reliable specific heritability estimates or if the responses are for different environments, average heritabilities may be substituted.
3. Cytogenetics . Cytological analyses by Maria et al . (1995, 1998a,b) on the karyotypes of E. guineensis and E. oleifera confirmed that both species have 2 n =32 chromosomes and had similar chromosome arm lengths for the three chromosome groups. These factors might explain the crossability of the two species and the partial fertility of their hybrids (Hardon, 1986). Genomic in situ hybridization (GISH), in which total genomic DNA is used as a probe for hybridizing to chromosome spreads, has been used to separate the parental chromosomes in oleifera × guineensis hybrids and would be useful in backcross breeding programs. There are two estimates of the oil palm genome size: Jones et al. (1982) estimated 1900 megabase pairs using Fuelgen microdensitometry while Rival et al. (1997) estimated 3400 using flow cytometry, the discrepancy attributed to the different methods used.