Nutrition: Results
[addw2p name=”oilPalmNutrition”]
B requirements of the oil palm
The B contents of the canopy and stem increased rapidly from 20 to 82 month after planting (Table 1). These increases were mainly due to palm growth since their B concentrations were relatively constant over the period of measurements. Root B content was highest (44 g ha-1) at 37 months after planting but it quickly stabilized thereafter at an average of 34 g ha-1 (Table 1). The B requirements for FFB yields and production of male inflorescences between the sampling intervals increased exponentially before reaching a plateau at 71 months after planting (Table 1), again due to biomass increments rather than B concentrations in the palm components.
The cumulative B contents for each palm component followed a non-linear regression model (Table 2). The cumulative B contents for canopy and stem fitted well to modified exponential models whereas those for roots and FFB yields to hyperbolic models. The latter implied that the B requirements were relatively constant after their maximum has been attained. The r-squares for all the equations exceeded 0.78.

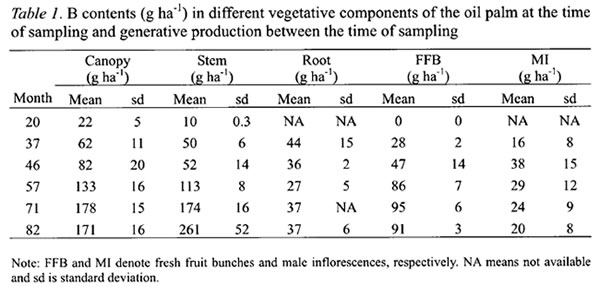
The annual B requirements of oil palm followed a sigmoidal curve with rapid increase from the second year after planting, reaching a maximum of 343 g B ha-1 yr-1 in the sixth year (Figure 1). The annual B requirement then declined to about 220 g B ha-1 yr-1 in the eighth year before stabilizing at 198 g B ha-1 yr-1 from the twelfth year. The latter was based on the estimated annual B requirement of the 16 years old palms at steady state condition in the second experiment. The highest B requirement of oil palm came from its developing canopy, which peaked at the fifth year after planting when the canopies of neighboring palms were fully overlapped (“closed”) and competition began to set in. At the peak, the B requirement for the canopy accounted for more than 40 % of the total B requirement of oil palm.
The B requirement for stem development was also the highest in the sixth year at 67 g B ha-1 yr-1 before declining sharply as palm competition intensified and growth rate slowed down (Figure 1). In contrast, the B requirement for roots was not only the lowest but also attained its peak earlier at the third year after planting (Figure 1). Thereafter, it required about 9 g B ha-1 yr-1 to sustain root turnover or self pruning of roots.
The B requirement for FFB yield was the second largest among the palm components and account for about 33 % of the annual B requirement at the sixth year and increasing to 37 % as the B needs for stem decreased sharply (Figure 1).
B distribution within the oil palm
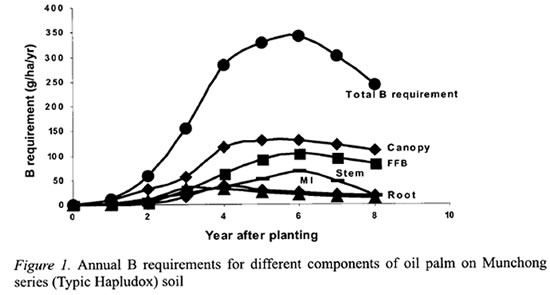
The detailed B distribution in the oil palm was studied in the second experiment where the palms were 16 years old and in steady state condition (Tables 3 and 4). The cabbage, which composes mainly meristemic cells, had the highest concentration of B at 12.3 mg kg-1 (Table 3). This was followed by the leaflets which were at the tail-end of the transpiration stream. Interestingly, the petiole, which supports the rachis and leaflets, and the petiole base, which are attached to the stem after the leaf is pruned off, had higher B concentrations than the rachis or stem. The stem and rachis had similar B concentrations of 3.3 and 3.6 mg kg-1, respectively. Although the stem B concentration was low, it has accumulated a large quantity of B by the 16th year at 404 g ha-1. The roots had the lowest B concentration of 1.4 mg kg-1.
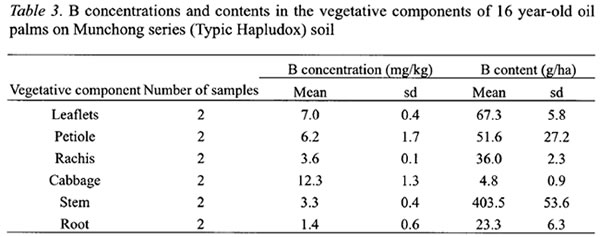
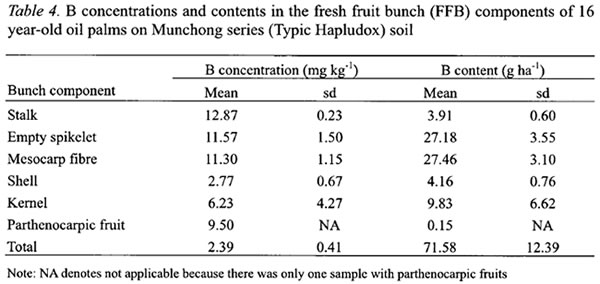
In FFB, larger B concentrations were found in the stalk, empty spikelets and mesocarp fibre (Table 4). These components have more rapid transpiration rate particularly during fruits development and oil formation stages (Jeje et al ., 1978). B concentration in the kernel at 6.23 mg kg-1was 2.25 times more than the shell (Table 4). The stalk and empty spikelets, which are the main components of empty fruit bunches after the milling process, contained about 43 % of the B in FFB.
A detailed analysis of the B distribution in the canopy showed that the B concentrations seemed to be relatively constant from the spear leaves to leaf number 32 before decreasing in the older leaves (Table 5). In terms of B content, there was an initial increase from leaf 1 to leaf 9 due to increasing frond dry weight. The B contents between leaf 9 and leaf 32 were similar at an average 26.3 g B per frond. The B contents then declined linearly between leaf 33 and leaf 48. This decrease was mainly due to lower dry weights and B concentrations of both petiole and rachis (Table 6). There was no clear trend in B concentrations in the leaflets.

B distribution in an oil palm plantation
The B distribution in the various components of one hectare of mature oil palms at stead state indicated that the largest store of B was in the stem at 404 g ha-1 followed by the canopy at 155 g ha-1 (Figure 2). The ground vegetation of mainly grasses and ferns contained very low B at only 6 g ha-1.
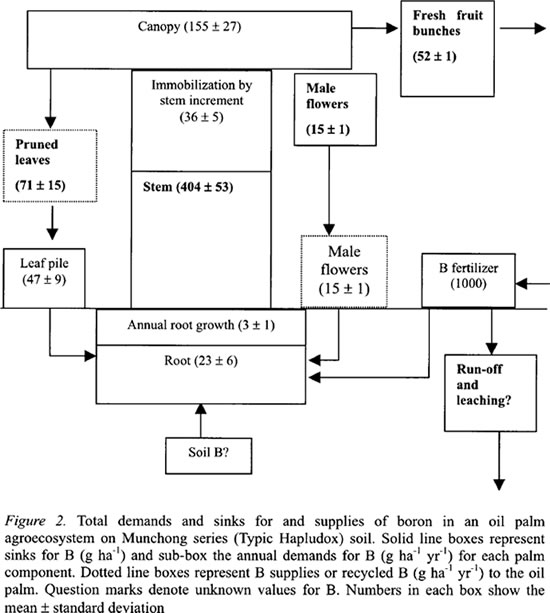
In terms of B cycling, the pruned leaves would return 71 g B ha-1 yr-1 to the agroecosystem and the decaying male inflorescences about 15 g B ha-1 yr-1. The pruned leaves were neatly stacked in leaf piles in the area between the palm rows called the inter-rows. The latter area stored about 47 kg B ha-1, which was 66 % of the annual B in the pruned leaves. The lower B value in the leaf pile might be attributed to the high decomposition rate of oil palm leaf where most of it would decay within six months.
In the oil palm agroecosystem, the FFB yields are transported to the oil palm mill for processing, and this component contained 52 g B ha-1 yr-1. The oil palm also immobilized about 36 g B ha-1 yr-1 mainly for stem growth.