Nursery: Set-up
The decision to have a single stage or double stage nursery will be a matter of personal choice depending on the specific situation encountered e.g. a double stage nursery is advisable for a large scale planting (>500 ha).
The Single Stage Nursery
This system utilises only large polybags and the germinated seeds are planted directly into the bags in the same manner as planting out in the pre-nursery polybags as described in section on planting germinated seeds into the large polybags.
Some of the advantages of this system are:-
-
once the seed has been planted, there is no further movement until field planting so that the root system is not disturbed and therefore establishment and growth is faster.
-
one can dispense with all the requirements in terms of layout, equipment and labour of the pre-nursery.
However this system has a number of disadvantages: –
-
it is necessary to have the full nursery infrastructure ready from the initial seed delivery.
-
it requires greater volume of water and additional engine fuel, wear and tear for the first two or three months.
- it also requires more soil since all the culling is done at the large polybag stage.
-
it is more difficult to observe and supervise when the seedlings are spread over a large area.
-
there will be no space for receipt of the following year’s seed delivery in case of delays in any one year’s planting programme unless the nursery is enlarged.
-
culling and seedling replacement is cumbersome.
-
it is not advisable for large scale planting which requires a very large area to prepare and also involves high cost.
-
difficulty in providing proper shading to the germinated seedlings at planting.
The Double Stage Nursery
The double stage nursey system involves planting of the germinated seeds in small pre-nursery polybags packed closely together in a very small area for the first two to three months. The seedlings are then planted out in the large polybag nursery where they remain for a further 7-9 months before field planting. This system has a number of advantages over the single stage nursery such as:
-
only a small section of nursery is required for the first 2 to 3 months.
-
less irrigation required for the first 2 to 3 months.
-
easier to observe seedlings and supervise nursery work during the critical early stage (2 to 3 months after planting).
-
culling can be carried out very quickly and easily at the first stage before planting into large bags.
-
the double stage nursery also has some disadvantages namely:
-
an “extra” operation is created which is very labour intensive.
-
possible slower growth compared to the single stage especially with poor shade management.
-
poor transplanting technique from small to large polybags could give rise to severe transplanting shock.
NURSERY SET-UP (Double Stage Nursery)
Since the double stage nursery is commonly adopted in most of the estates, only this type of nursery is described.
The Pre-Nursery Stage
Polybag filling and placement
Polybags must be filled with soil up to the bag rim and placed in the nursery beds at least four weeks before the planting date to allow settling, topping up with soil and pre-planting irrigation.
Only the best topsoil available should be used in any nursery. The soil must be free draining, friable, sandy clay loam (eg. Rengam, Serdang, Bungor) and free from contaminants (chemicals etc.). Rock phosphate (RP) (@ 10 kg/1000 small polybags of 15 cm (6 in) x 21 cm (9 in) x 250 gauge) must be premixed into the soil prior to filling to ensure adequate P availability.
The polybags should be turned inside out before filling so that they sit upright. Hoppers or funnels may be used to facilitate filling. The filled polybags should be arranged in beds of 10 bags width and of a convenient length. The beds should have wooden side frames to prevent bags from toppling over. The arrangement of polybags at pre-nursery stage is shown in photo 1.

Shading
At this critical stage it is necessary to shade the plants either with palm fronds on frame, or with shade netting of 70% shade. If oil palm fronds are used as shade it is necessary to pre-spray the fronds with a pesticide to ensure that they are not instrumental in introducing any pests or diseases. The shade should be gradually reduced after the first month and should be totally removed by the end of the second month. The shading in the pre-nursery stage using oil palm fronds and netting is shown in photos 2 and 3 respectively.

Planting germinated seeds into the polybags
It is essential to ensure that polybags are well watered just prior to the commencement of planting. Planting of germinated seeds should be completed as soon as possible, preferably not later than 1 day after receipt.
Care must be taken to ensure that the workers can differentiate between the plumule (shoot) and the radicle (root) to avoid planting the seeds upside down. The seed should be planted at 1 cm depth with the radicle pointing down and lightly covered with soil. Planting holes can be made with a stick.
Where the germinated seeds are consigned as identified families and labelled accordingly by the seed producer (as for AA DxP seeds), they are to be planted according to the same families and properly labelled right to the main nursery. This is to facilitate nursery culling, as the seedlings from the each family would be more uniform and the off-types readily spotted. However in the field planting, the seedlings are to be planted as mixed families.
Once the planting is completed, the identification tag must be stapled to the edge of polybag.
The Main Nursery
Polybag filling
Again, only the best topsoil is to be used in the large polybags . Prior to polybag filling, a basal application of 100g RP/polybag must be thoroughly mixed with the soil before filling.
Filling of the polybags should commence at least a month prior to the transplanting. Bags should be filled to the rim and set aside for a few weeks to allow for settling and soil topping up where necessary.
The large polybag of 38 cm (15in) x 45 cm (18in) x 500 gauges should be turned inside out before filling, to give the filled bag a stable and level base.
Hopper or funnels may be used for more efficient filling especially in large-scale nurseries.
Spacing of bags
Spacing of polybags in the nursery is done to minimise light competition between palms and for easy access to the palms for pest and disease control, weeding and manuring. The spacing and placement of seedlings at main nursery stage is shown in photo 4. The spacing of seedlings depends on the duration seedlings are expected to be kept in the nursery. The recommended spacing for various seedling ages at field planting is given in Table 1.
|
Table 1: Recommended polybag spacing in the nursery for various seedling ages at field planting
|
||
|
Age of seedling at field planting (months)
|
Triangular spacing of polybags in the nursery
|
|
|
(m)
|
(ft)
|
|
|
09-11
|
0.75
|
2.5
|
|
11-13
|
0.91
|
3
|
|
13-18
|
1.25
|
4
|

Transplanting from pre-nursery into large polybags (main nursery)
Prior to transplanting the seedlings to the large polybags, workers should prepare the planting holes within the bags. These holes can be prepared using core-borers. The pre-nursery seedlings should be given a thorough watering before the transplantings. To minimize disturbance of the roots, the bags should be cut open with a small blade and the seedling removed gently without disturbing the soil. The detached seedling together with the soil is placed in the planting hole of the large polybag, and the soil gently but firmly compacted around the root mass. Care must be taken to ensure that the planting is level with the pre-nursery soil level. A good watering should follow immediately. Mulching of the soil surface should be done with oil palm kernel shells.
Nursery: Ordering of Seeds
Placing of Orders
Seeds are usually ordered well in advance of requirement. It is normal to place tentative orders at least one year before delivery subject to confirmation at a date closer to delivery. Delivery must be spread out to provide time for planting out at the optimum age.
Source of Seeds
All germinated seeds, or any other planting materials, should only be purchased from reputable suppliers. Wherever possible seeds should be purchased direct from SIRIM registered producers to be ensured of legitimate and good quality seeds .
Quantity to Order
In determining the quantity to be ordered , the culling rates and various losses (in the nursery and field) will have to be considered in addition to the total palms required in the field. Based on general experience, 175 germinated seeds (27% more) per hectare are sufficient if the field stand is 138 palms per hectare. For a stand of 148 palms/ha, the requirement would be 186 germinated seeds.
Nursery: Site Selection and Preparation
SITE SELECTION
The selection of the area for a nursery is critical and it should be sited as centrally as possible to the field(s) to be planted. In addition, the following should also be considered:
Topography (Terrain)
The selected area should be flat to gently undulating with slopes between 0 and 30 and preferably, with a reliable/permanent source of water supply for irrigation purpose.
Water Supply
Water requirements (quality and quantity) must be determined prior to starting site preparation. The easiest source of water is where there is a large natural pond or lake whereby all that is required is to place an intake pipe to connect with a pump unit. A back-up system should be considered, particularly in isolated areas or areas of lower or unreliable rainfall.
Drainage
The site chosen should not be prone to flooding, which will damage seedlings and buildings (stores).
Area
To achieve good growing conditions with minimal risk of etiolation, a main nursery planting density of 13,800 polybags per ha with 0.91m(3ft) x 0.91m(3ft) D planting is recommended excluding allowance for accessibility. The spacing should be increased by another 0.15m (0.5ft) if the seedlings are anticipated to be kept in the nursery for longer than 12 months.
Accessibility and Nursery Roads
Roads within the nursery and their alignments will need to be carefully planned and laid out depending on the placement distance of the polybags and the type of irrigation to be utilised. Access roads to the nursery should be sufficiently wide to allow vehicles to manoeuvre during peak planting periods to facilitate supervision and movement of materials.
SITE PREPARATION
The preparation of the area for a nursery is important to allow optimum seedling growth, maintenance of nursery site, unimpeded access and to provide hygienic conditions for plant growth. Four main activities are involved in preparing a site for nursery, namely nursery design, clearing, fencing and lining.
Nursery Design
A well-designed nursery allows for access of many vehicles during evacuation of seedlings for field planting especially for large-scale plantings. This objective can be achieved through the drawing up of a plan to show all paths, roads and irrigation points. An example is shown in Figure 1.

Clearing
With the boundaries determined, felling and clearing should be carried out at least 2 months before the arrival of the seeds. Once clearing is completed, proceed to fence the area, fill the polybags and install the irrigation system.
Fencing
The major types of fences utilised for nurseries are the conventional barbed wire fence and the electric fence.
The Conventional Fence
The specifications for the conventional fence depend on the species of animals that it is required to keep out. For example, a four-strand barbed wire fence, with wires spaced at 0.3, 0.6, 0.9 and 1.2 m from ground should be adequate to control cattle and goats.
Electric Fencing
Where there is threat of wild mammalian pests, an electrical fence is possibly the best form of defence. Wires of 250-300 kg breaking strain have been found to be the most suitable. The heights at which the electrified wires are placed above ground level are critical, being dependent on the mammalian pests to be kept out, e.g. 10 cm to repel porcupines.
Lining
Lining is carried out to space the polybags evenly in the nursery, so that seedlings have good and uniform access to sunlight and to achieve the most cost effective irrigation system .
Polybags are lined at 0.91m (3 ft) x 0.91m (3 ft) triangular spacing to give each seedling the optimum growth space. All seedling rows must be straight along the axis at 60o to each other and parallel to irrigation lines.
In the Sumisansui irrigation system, MK II tubes are laid down between the polybags at every 4 rows, a 1 m wide path is provided at every 8 rows for easy access. When the seedlings are about 7 – 8 months old, additional MK II tubes are added between the existing MK II tubes to give a final layout of 1 tube for every 2-seedling row. This is illustrated in Figure 2.

Oil Palm: Nursery
NURSERY PRACTICES FOR PRODUCTION OF SUPERIOR OIL PALMPLANTING MATERIALS
Introduction
Site selection and preparation
Ordering of seeds
Nursery set-up
Nursery maintenance
Summary of recommendations
The oil palm is a very precocious crop, which comes into maturity at 25 months or earlier from field planting, with peak yield realized at four or more years thereafter.
As early bearing and high yields in the field are mainly dependent on production of uniformly good and healthy seedlings from a nursery, it follows that good nursery management would be required to achieve the latter.
The production of superior oil palm planting materials is fully dependent on attention to details at all stages in the nursery management and this entails following closely, proven standards and procedures.
Reference
Heriansyah and Tan C.C. 2005. Nursery practices for production of superior oil palm planting materials. The Planter. Incorporated Society of Planters, Kuala Lumpur 81 (948) : 159-171
Note: The full list of references quoted in this article is available from the above paper.
Oil Palm: Replanting
Some pulverisation techniques of clearing old oil palms for replanting
- Chip and burn (open burning has been banned)
- Chip and windrow
- Chip-windrow and pulverize when palm chips have partially decayed
- Poison and under-plant
Why do we need improved methods of clearing old palms for replanting?
- The ban on open burning resulted in very high breeding of rhinoceros beetles, now the most serious pest in immature and young mature palms in Malaysia
- Liau et al . (1991) reported crop loss of up to 40% in the first year of harvest at 50% beetle damage incidence
- Chung et al . (1999) reported that severe beetle damage could result in 92% loss of the first 12 months’ crop
- Windrowed palm residues hindering field operations
- Nutrients in windrowed palm residues not readily available to replanted palms until the palm roots have reached the windrows
- Windrowed palm residues harbouring and breeding rats
1. Telok Sengat Estate method (tractor-mulcher)
-
The entire palm is pulverized into small pieces and spread widely over the field and the root mass around the bole dug up during land clearing .
-
Tractor-Mulcher pulverising 40-day old chips
-
Tractor ploughing up missed and partially pulverised chips
-
Second round pulverisation after ploughing
-
Field condition after 2 rounds of pulverisation and a round of ploughing in between

2. KLK method (excavator-mulcher)
-
Excavator fell and chip palm and spread chips into single-palm heaps
-
Excavator-Mulcher™ pulverizing 2-month old palm chips

Three techniques for pulverising the standing and newly felled palms
Technique 1. The EnviroMulcher™
-
Cylindrical steel drum with 111 pieces tungsten carbide knives and mounted at the excavator’s boom. It is driven by the hydraulic system of the excavator
-
Pulverizing a fallen trunk including the crown
-
A supporting excavator digs up root mass and stacks fronds

Technique 2. The MountainGoat™
-
Powered by a 750hp caterpillar engine and supported by 2 excavators
-
Two excavators fell and line palms for a MountainGoat™
-
The same excavators also dig up root mass
-
MountainGoat™ chipper drum with 10 knives

Technique 3. The Beaver™
-
Powered by a 300 hp caterpillar engine
-
Severing root mass with blade
-
Beaver felling palm
-
Rotating drum with 20 pieces self-sharpening flails pulverizing felled palms
-
Pulverizing a felled palm
-
Mulching a newly planted palm
-
Mulching newly replanted palm circle with freshly pulverised chips produced by EnviroMulcher™ improves palm growth significantly

Conclusions
The major problems associated with the conventional methods of land clearing could be overcome or minimized by pulverizing and spreading the palm biomass widely over the field during land clearing prior to the planting of palms and legume cover crops
References
Ooi L.H. and Heriansyah 2005. Palm pulverization in sustainable oil palms replanting. Plant Production Science 8 (3) : 345-348.
Ooi, L.H., Kodiappan, P. and Gunarajan, M. 2004. Some pulverisation techniques of clearing old palms for replanting. Proc. National Seminar on Replant or Perish. Incorporated Society of Planters, Kuala Lumpur: 47-63.
Chung, G. F., Sim, S. C. & Balasubramaniam, R. (1999). Effects of pest damage during immature phase on the early yields of oil palm. In 1999 PIPOC Proceedings: Emerging Technologies and Opportunities in The Next Millennium. Palm Oil Research Institute of Malaysia, Kuala Lumpur, pp 454-476.
Liau, S. S. & Ahmad Alwi. (1991). The control of Oryctes rhinoceros by clean clearing and its effect on early yield in palm-to-palm replants. In: 1991 PORIM Int. Palm Oil Conf. pp 396-403.
(Movies and photos contributed by Ooi Ling Hoak, unpublished)
Plant Breeding: Future Prospects
The palm oil industry will continue to grow for the following reasons:
-
growing world population and the still low per capita consumption of oil
-
high productivity of the crop and low cost of production
-
versatility of palm oil and palm kernel oil in oleochemical and industrial uses
-
being a renewable resource, the edge over mineral oil in the increasingly environmental conscious world
Biofuel from vegetable oil is currently used in the countries of the European Union. Similarly, palm diesel will soon make its scene in the palm oil countries e.g. Malaysia, for economic and strategic reasons, although crude palm oil has been traditionally used to power vehicles in plantations in West Africa.
The big stake in the crop in many emerging economies demands continuing funding for research and development (R&D). The need for increasing investment in R&D is even more crucial for higher cost producers e.g. Malaysia, because of the tremendous biotechnological advancements made in competing annual oil crops such as rapeseed and soybean and the subsidies provided which erode the competitive edge and profitability of palm oil. Cultivar improvement is strategically an important if not the primary objective of most crop R&D efforts. Oil palm breeders will have to be able to respond readily to the changing market needs. The first need is to improve crop productivity, and the trend is towards precision farming to achieve this objective. Specific cultivars adapted to certain environments and plantation management requirements are the first prerequisites. The production of definite hybrids (single cross, biclonal) and clones is directed towards this end. Cultivars must be management-friendly in terms of ease of mechanization of harvesting and implementation of other plantation operations. The second is to extend the versatility of the crop in terms of added value products (Murphy 2000; Murphy and Peterson 2000) especially when its high productivity confers an existing comparative advantage. A good example is the production of polyhydroxy butyrate (PHB) for biodegradable thermoplastic production, a recently initiated collaborative project in Malaysia. Others would include production of ß-carotene for vitamin A and natural dye, tocotrienol (vitamin E), industrial fatty acids such as petroslenic acid, erucic acid, and ricinolic for specialty plastic and lubricant production. Most of these objectives will have to be achieved via biotehnological means. It can be argued that the oil palm is not the most appropriate crop for biotechnological genetic modifications because of its perennial nature and the lag time involved (Corley 2000; Hardon and Corley, 2000; Tinker 2000). However, the very rapid development of biotechnological techniques and tools e.g. microarray or DNA chip technology, bioinformatics, and the revelation of much genetic collinearity or synteny (similarity or identity of genes) between crops suggest the genes, techniques and information developed in the cereal crops can be used in oil palm to reduce the development time. And if the transgenes have been strategically incorporated into advanced breeding parents or proven clones, the response time in production of new hybrids to meet market changes would be considerably hastened. Indeed, oil palm breeders will have a challenging but interesting time ahead.
Plant Breeding: Improvement Objectives
A. Yield Potential
The oil palm is the highest yielding oil crop (Corley 1985; Robbelen 1990), its highest annual recorded oil yield of 10 t/ha is 2-5 times those of annual oilseed crops such as soybean and rapeseed. Yield improvement in the oil palm plantations for the past 50 years has been attributed to 70% cultivar improvement and 30% improvement in agronomic practices (Davidson 1993). Breeding progress in the Deli dura planting materials was about 15% per generation. (A breeding generation in oil palm requires 8-10 years; three years to bearing, four to six years of yield measurement, and one year of hybridization). With the switch-over to the tenera planting material there has been at least 30% improvement in oil yield mainly through better oil content from the thicker mesocarp (Hardon et al. 1987). Since then there have been two generations of improved tenera hybrid planting materials and oil yields have improved from 4.9 t/ha to 8.9 t/ha but this could not be attributed entirely to breeding as the materials were planted at different times and locations and perhaps with different agronomic inputs (Lee and Toh 1992).
Current planting materials derived from mixed hybrids thus possess high yield potentials. The theoretical yield potential based on crop physiological computations is 17 t/ha oil from 45 t of fresh fruit bunches (Corley 1985). Oil yields exceeding 12 t/ha, fresh fruit bunch yields of 45 t/ha, and oil content in bunch of 35% have been reported in experimental plantings, although not necessarily jointly. Nevertheless they indicate that the prospect of significant quantum increase in yield potential is rather limited, at least in the near term. For larger quantum yield improvement drastic changes with the crop architectural design (plant form and planting pattern), cultivation and harvesting methods would be needed.
Increase in crop yield potential in other crops has been achieved through the combination of high biomass production and high harvest index, the former through high density planting (Evans and Fischer 1999). With current genotypes and cultivation system (uneven terrain plantings and high dependence on manual operations particularly for harvesting) plant densities of higher than 138 to 160 palms/ha currently practiced would seem inadvisable. However, proposals and efforts are underway to breed for high efficiency palms with more efficient light capture, higher photosynthetic rate, higher leaf expansion ratio, higher leaf area ratio, better conversion efficiency of energy captured to dry matter, reduced respiratory loss and improved harvest index (Breure and Corley 1983; Squire 1984; Breure 1985, 1986; Henson 1992; Smith, 1993). Many of these contentions are based on inferences drawn from measurements made on individual palms in mixed stands and on correlations based on small sample sizes which were prone to auto-correlations or statistical dependencies because most of the physiological parameters were derived variables in which total dry matter or yield constituted a major component (Rajanaidu and Zakri 1988; Chang and Rao 1989). Proof of the value of these physiological or ideotype breeding traits (Rasmusson 1991) would have to be obtained from replications of such attributed palms within plots and between plots in experimental plantings. These are as yet not forthcoming. In cereals, higher yield potential has been achieved through higher biomass and/or better harvest index using dwarfing genes (Peng et al. 1999; Reynolds et al. 1999) while increased yield in maize has been attributed to tolerance to high density planting (Tollenaar and Wu 1999). The Dumpy semi-dwarfing gene(s) in oil palm did not contribute to better harvest index (Soh et al. 1981). Despite this, slight increases in harvest index and planting densities are conceivable with new genotypes in the breeding pipeline (Rajanaidu et al. 2000).
While the concept of genetic yield potential is helpful in achieving the physiological limit, the concept of harvestable yield is important for a perennial tree crop where the actual harvesting (cutting) operation (Plates 1B, 1C) is manual with little prospect of a feasible and cost effective mechanized alternative. Harvestable or recoverable yield relates to ease of harvesting, harvesting at optimal fruit ripeness and minimal loss of ripe fruits. Dwarf palms with longer bunch stalks will facilitate harvesting, manual or mechanical (Soh et al. 1994b). Palms with ripe fruits exhibiting a distinct color change will ensure harvesting at optimum ripeness while those with non-abscising fruit habit will minimize loose ripe fruit loss. Such programs have been proposed using breeding or genetic engineering approaches (Osborne et al. 1992; Rao 1998) as these traits are found in less advanced breeding or germplasm materials.
 Plate I. B) Four year old oil palm harvested with a chisel. |
 Plate I. C) A 20 year old oil palm harvested with sickle attached to aluminum pole. |
B. Adaptability
Much of the reported planting material improvement is likely to have been through adaptability to site limitations. This involves moving the site yield potential (Tinker 1984) towards the genetic yield potential by circumventing site limiting factors e.g. soil, moisture, terrain, rather than through improvement in genetic yield potential per se. However, genotype × environment (GxE) interaction has not been generally considered to be a serious factor (Rosenquist 1982; Cochard et al. 1993). The presence or absence of GxE depends on the specific genotypes and specific environments tested (Corley et al . 1993). The misperception that GxE is not important could be attributed to the use of the genetically variable commercial sources of planting materials as the genotypes (Chan et al. 1986; Rajanaidu et al. 1986); the use of related progenies in different environments (Cochard et al. 1993) or the use of related progenies in similar environments (Rosenquist 1982; Lee and Rajanaidu 1999). Results of other experiments and analyses (Obisesan and Fatunla 1983; Obesisan and Parimoo 1985; Ong et al. 1986; Corley et al . 1993) have detected the presence of GxE effects using genetically diverse progenies although their contributions (3-4%) to the total experimental variance for yield were still small (Rajanaidu et al., 1993; Rafii et al., 2000). Although purported stable progenies have been identified (Ong et al. 1986; Lee et al. 1988; Rafii et al. 2000) their exploitation as cultivars depends on the ability to reproduce these tenera hybrid progenies in large quantities, as only limited hybrid seeds can be produced from a pair of dura and pisifera parents. Most of these authors also did not indicate the characteristics that conferred stability in the progenies or attempt a biological explanation of the GxE effects (Caligari 1993; Lee and Rajanaidu 1999). Corley et al. (1993), using a simple analytical approach found that progenies with few bigger bunches rather than many smaller bunches tended to yield poorly in stressful environments because the abortion of a single large bunch would result in a considerable loss in yield. Plasticity of bunch weight was also found to vary between progenies.
In terms of selection, an attractive approach used in animal breeding to handle GxE effects is to treat a trait in different environments as separate but genetically correlated traits (Falconer and Mackay 1996). They can then be incorporated into a selection index to predict selection response for the trait in the second environment as a correlated response to selection for the trait in the first environment (White and Hodge 1989; Yamada 1993; Soh 1999).
With the advent of near true hybrid cultivars from inbred parents or cloned parents and clones, GxE effects with respect to location, spacing, fertilizer, and other factors will likely assume increased importance (Lee and Donough1993; Corley et al. 1995; Soh et al. 1995) and may warrant consideration in the development of cultivars suited for different situations, in line with the trend towards precision farming or precision plantation practices (Chew 1998).
C. Oil Quality
The oils produced by the oil palm are very versatile in their uses in the food and oleochemical industries. Despite this, palm oil has yet to penetrate the cooking and salad oil markets of the temperate countries. Its large saturated fatty acid component (Table 1) makes the oil with a melting point of 36ºC (Yusoff 2000) solidify at colder temperatures. Because of this and the fact that it is a palm-derived oil, consumers have incorrectly lumped it with coconut oil as a saturated fat unhealthy for human consumption leading to a predisposition to heart disease. This is despite the fact that palm oil contains about 50% unsaturated fatty acids and that the saturated component is mainly palmitic acid which Khosla and Sundram (1996) contended to be neutral in its cholestrolemic behavior. Palm oil also contains anti-oxidants in its carotene and tocopherol/tocotrienol contents which appear to have anti-carcinogenic properties. It can also be used directly, i.e. without hydrogenation, for margarine without the production of the unhealthy trans fatty acids.
Table 1 . Variability of fatty acid composition and iodine value in oil palm populations. Source: Arasu 1985; Rajanaidu 1990a; Rajanaidu et al . 2000.
|
Fatty acid |
Content (%) |
||||
|
Nigerian |
PORIMz |
IRHOy |
E. oleifera |
E. oleifera ×
|
|
|
C14:0 |
0.3-3.1 |
0.9-1.5 |
0.3-1.6 |
0.1-0.3 |
0.1-0.5 |
|
C16:0 |
37.4-46.6 |
41.8-46.8 |
34.7-50.1 |
14.4-24.2 |
22.4-44.7 |
|
C18:0 |
3.8 -14.7 |
4.2-5.1 |
3.1-8.8 |
0.6-2.2 |
1.4-4.9 |
|
C18:1 |
33.0 -55.9 |
37.3-40.8 |
32.0-46.0 |
55.8-67.0 |
36.9-60.1 |
|
C18:2 |
5.4 -15.8 |
9.1-11.0 |
10.0-16.0 |
6.0-22.5 |
8.3-16.8 |
|
–
|
Iodine value |
||||
|
43.8-69.8 |
51.0-55.3 |
– |
67.4-91.9 |
– |
|
zPalm Oil Research Institute of Malaysia.
yInstitut de Recherches pour les Huiles et Oleaigneux
Nevertheless, end-user and consumer bias has pressured breeders to breed for a more liquid palm oil. The ideal palm oil should have the following fatty acid composition based on the American Heart Association’s (1990) recommendated fat intake: total energy requirement derived from dietary fat intake should not exceed 30%; one third from saturates [lauric (C12:0), myristic (C14:0), palmitic (C16:0), stearic (C18:0)] which are hypercholestrolemic; one third from polyunsaturate [linoleic (C18:2)] which is hypocholestrolemic; and one third from monounsaturate [oleic (C18:1)] which is neutral cholestrolemic. In this respect, palm oil is excessive in saturates and deficient in polyunsaturates.
Advanced oil palm breeding populations have low genetic variability for unsaturated fatty acid contents, generally expressed as iodine value. The situation is better in the West African semi-wild prospected materials but the highest values can be found in the American oil palm, E. oleifera , collections. Palm Oil Research Institute (PORIM), currently known as Malaysian Palm Oil Board (MPOB), recommended elevating the iodine value to about 70 by breeding to put palm oil on a competitive basis with olive oil (Task Force 1985). To do this without sacrificing the current progress in yield and agronomic attributes using selections from the collected E. guineensis materials would involve 4-5 generations of backcross breeding to the advanced breeding materials. With the E. oleifera materials, more backcross generations are envisaged because of interspecific hybrid infertility (Hardon 1986, Sharma 2000). The same would apply to breeding for higher carotene and tocopherol/tocotrienol contents. Reservations to this approach have been expressed (Hardon and Corley 2000). Because of the long lag breeding time (20-30 years), by the time the cultivar is developed, the market may have changed and the perceived premiums disappeared. Genetic and agronomic manipulations are more expedient with annual crops, which can respond readily to market changes. An oil quality genetic improvement program also requires a large separate effort, which may be at the expense of the main yield improvement program through dilution of effort and selection pressure. It may be more expedient to concentrate on yield improvement and achieve higher quality component production from more oil produced and using chemical and bioprocessing technological advances.
D. Stress Tolerance
Being a profitable and easily grown crop, oil palm plantings have expanded into sub- optimal and marginal areas (e.g. dry sandy areas, podsols, peat, highlands). In these areas biotic (disease and pest) and abiotic (water, temperature, nutrient) stresses, which impede or reduce production, are likely to be encountered.
1. Biotic. Basal stem rot caused by the basidiomycete fungus Ganoderma boninense is the only disease warranting consideration in resistance breeding in the Far East. It used to be a malady of older palms planted on former coconut or oil palm areas with high water tables but reports of its occurrence on younger plantings in inland areas have become more frequent (Ariffin, 2000). Progress in resistance breeding had been hampered by the lack of an efficient screening technique (Khairudin et al. 1993; Ariffin et al. 1995). No resistant or tolerant genotype has been found to date with the preliminary screening technique developed (Ariffin, 2000). In a very recent paper (De Franqueville et al 2001) parental, progeny and clonal differences in resistance/susceptibility were demonstrated in palms planted on Ganoderma infested soil. Ganoderma disease control is the subject of an international cooperative effort coordinated by CAB International (1998) in which resistance control is an important objective.
Because a reasonably efficient nursery screening technique is available, tolerance to Fusarium (Fusarium oxysporum f. sp. elaedis) vascular wilt forms an integral part of the breeding programs in West Africa (De Franqueville and Renard 1990). Lethal bud rot debilitates oil palm in Latin America. Any proposal to breed for tolerance to this disease would be premature as the pathogenic cause of the disease is still in contention (Ariffin 2000).
Crown disease, a physiological affliction of young 2-3 year old oil palms in the Far East, causes bending and twisting of the young fronds, which can result in loss in early yields when severe and prolonged. The Deli dura material appears to be more susceptible. The disorder is caused by a recessive gene, the expression of which is masked by an epistatic gene conferring incomplete penetrance (Blaak 1970). The disorder can be bred out by discarding families with any susceptible progeny.
2. Abiotic. Oil palms in West Africa experience reasonably long periods of drought and hence West African commercial planting materials tend to be more drought tolerant because they have been bred under such conditions. There are also drought tolerance breeding programs (Houssou et al. 1987; Okwuagwu and Ataga 1999). The semi-wild collections obtained by MPOB from the drier regions of Nigeria would include drought tolerant genotypes. Palms introduced from the Bamenda Highlands of Cameroun (Blaak 1967) are presumed to possess genes for cold tolerance although their likely precocious flowering behavior under lowland conditions have been touted.
Manifestations of Mg deficiency occur on oil palms planted in sandy areas in some parts of Papua New Guinea and Indonesia. Tolerance to Mg deficiency is an objective of the breeding program in Papua New Guinea (Breure et al. 1986). Palms planted on deep peat tend to lodge. Dwarf or smaller palms would circumvent this problem. Peat plantings are also prone to micronutrient deficiencies, particularly Cu, Zn, and B. As these deficiencies can be corrected easily with micronutrient applications, a breeding approach is unnecessary.
Plant Breeding: Breeding Techniques
A. Floral Biology
The oil palm is cross-pollinated and monecious bearing a staminate or pistillate inflorescence in each subtending leaf or frond (Plates 1D, 1E). Inflorescences, which are compound spikes, begin to appear on young palms when they are about 27 months old from seed sowing, which is also the same length of time from inflorescence initiation to its emergence from the sheath in mature palms (Henson 1998). Sex differentiation occurs at the 14th month from inflorescence initiation. Stress conditions at this time will induce staminate inflorescence while at the 23rd month will result in inflorescence abortion and non-appearance of inflorescence on the subtending frond. There should be potentially two to three inflorescences produced per month corresponding to the frond production rate. Pollen viability and stigma receptivity last about 5-6 days and 3-4 days in the field respectively. The main pollination agent is the pollinating weevil, Elaedobius camerounicus, which occurs naturally in West Africa and Latin America but was only introduced to the Far East in the early 1980s. Controlled-pollination for breeding and commercial hybrid seed production involves the following protocol:
- Pistillate and staminate inflorescences are isolated 7-10 days before receptivity/anthesis using terelene/paper/canvas bags with clear plastic windows.
- Pollen can be stored as oven-dried pollen in the freezer for about 6 months or as freeze-dried pollen in vacuum-sealed ampoules in the freezer for about 24 months.
- Controlled-pollination is effected by puffing a 1 pollen: 10 to 20 talc mixture through a perforation in the plastic window (Plate 1F).
- The bag is removed after about a month and the fruits left to mature and ripen in about 5 months.
- The controlled-pollinated seed from the depulped ripe fruit needs to undergo 40-60 days heat treatment at 37º-38ºC and 17-19% moisture to break its dormancy before it can germinate when remoistened to 21-22% (Plate IIA). These are the requirements for germinating dura seeds. For tenera seeds the corresponding moisture requirements are 20-21% and 27-28% respectively. Germination of the shell-less pisifera and thin-shelled tenera seeds is erratic (Arasu 1970) and in vitro (embryo rescue) germination is sometimes needed.
 Plate I. D) Staminate inflorescence at anthesis, stage bagged inflorescence harvested for pollen collection. |
 Plate I. E) Receptive pistillate inflorescence. |
 Plate I. F) Pollination of bagged pistillate inflorescence. |
 Plate II. A) Germinated hybrid seedlings. |
B. Breeding Plans and Selection Methods
The oil palm being a cross-pollinated crop borrows a number of breeding methodologies developed in maize breeding such as recurrent selection and topcross testing, but being a perennial tree crop also shares many similarities with animal breeding in the selection techniques such as the importance of sire or pisifera testing, simultaneous multiple trait or index selection, and the close temporal and genetic correspondence of commercial hybrid production with each cycle of breeding.
1. Recurrent Selection. Major oil palm breeding programs adopt one of two basic schemes; the modified recurrent selection scheme and the modified reciprocal recurrent selection scheme.
Modified Recurrent Selection Scheme. This scheme is practiced by most programs in the Far East (Fig. 2). It evolved from the initial use of the Deli dura as the commercial planting material to its subsequent use as a maternal parent in commercial tenera hybrid seed production. Pisiferas, introduced or bred, and being female sterile, are initially selected based on their tenera-sib performance in the tenera × tenera/pisifera cross or based on proven lineage. They are then progeny-tested (top-crossed) with a sample of phenotypically selected Deli dura mother palms. Proven pisiferas and the phenotypically selected duras are then used in the commercial mixed tenera hybrid seed production. Parents used for further breeding in dura × dura and tenera × tenera/pisifera crosses are also phenotypically selected although progeny-tested parents are also sometimes included. As parent selection often involves family with individual palm selection, this scheme is sometimes referred to as the family and individual palm selection scheme (Rosenquist 1990). But as selected parents are interbred in each cycle, Soh (1990) preferred to refer to it as a form of recurrent selection (Allard 1960). This scheme emphasizes and exploits general combining ability (GCA) effects.
The main advantages of this scheme are that more recombinant crosses can be made and tested and in saving time, space and effort from need of extensive progeny-testings. The main disadvantage is that the parents chosen for further breeding and the dura mother palms for hybrid seed production usually have not been hybrid progeny-tested. The key assumption that the additive or GCA effects expressed within population crosses (determined by family and individual means or combining ability analyses) will be reflected in the inter-population hybrid crosses, may be untenable (Soh 1987b, 1999; Soh and Hor 2000). The scheme may also lead to undue prolonged reliance on the Deli as the source of dura mother palms.
Modified Reciprocal Recurrent Selection. This scheme (Fig. 3) practiced mainly in West Africa e.g. Ivory Coast, Nigeria (Sparnaaij, 1969; Meunier and Gascon, 1972) and more recently in Indonesia (Lubis et al 1990) is adapted from the reciprocal recurrent selection method developed in maize breeding (Comstock et al.1949). It exploits both GCA and specific combining ability (SCA) effects. The choice of this scheme presumably arose from the observations and results that crosses of the Deli duras with African teneras or pisiferas tended to give heterotic yields.
This attractive scheme has three apparent advantages. Firstly, the selfed parents selected for commercial hybrid production and for further breeding are both based on progeny-tests (from the dura and tenera grandparents) of the prospective commercial inter-population hybrids. Secondly, the scheme has two distinct phases. The “within hybrid improvement phase” allows shorter term commercial exploitation of the best test-cross and its improvement by recurrent selection within the selfs/sibs of the selected parents. The “recombinant phase” involving outcrosses, which is run in parallel similarly, allows accumulation of favorable alleles (additive and non-additive) and maintenance of genetic variability for sustained longer term improvement. Thirdly, the commercial hybrid can be reproduced using the duras and pisiferas from the selfs/sibs of the parents (which have been planted concurrently as the progeny- tests) once the inter-population hybrid test results are known (Jacqmard et al. 1981). Its main disadvantage is the large program size requirement. To produce three to four million hybrid seeds from the reproduction of the top 15% of the crosses, about 500 crosses and 180 parental selfs have to be planted over 600 ha and evaluated over 15 to 25 years (Soh 1999).
Breeding Progress. In the reciprocal recurrent selection scheme, the original cross from which subsequent breeding improvements are made can be reproduced to be the standard control cross for measurement of breeding progress achieved. For the first cycle of the reciprocal selection scheme, Gascon et al. (1988) reported a 18% oil yield improvement by selecting the top 2.8% of the 529 hybrid crosses tested for reproduction as commercial hybrid seeds. From early trial results of the second cycle (within hybrid improvement) hybrids, Nouy et al. (1990) predicted a 10-15% oil yield increase which was subsequently confirmed (Cochard et al. 1993). Lubis et al. (1990) suggested a 25% oil yield increase by selecting the top 8% of the 500 hybrids tested in the first cycle of the Indonesian Oil Palm Research Institutes’s (IOPRI) reciprocal recurrent selection program. Progeny-test results for the recombinant phase i.e. recombinant parents or wide crosses, are still unavailable. An important consideration for this phase is whether to use the selfs of the recombinant parents for the production of commercial hybrids which would result in higher within hybrid variability or to await another generation of selfs in order to produce more uniform hybrids. In the study of Durand-Gasselin et al. (1999) on 2, 3 and 4-way hybrid crosses, the increased within cross variability for production traits was minimal but these crosses involved related parents rather than outbred or recombinant parents. Also in this study, as reported earlier by Nouy et al. (1990), breeding progress was achieved mainly through exploitation of GCA effects although SCA effects might have played a small part.
It is difficult to estimate the breeding progress made by the modified recurrent selection scheme with confidence due to the lack of common link crosses across experiments and common base reference populations. Nevertheless, some indications can be obtained by piecing together some experimental results reported.
Hardon et al. (1987) estimated 15% oil yield progress per generation from breeding within the Deli dura but did not translate this in terms of improvement in the corresponding tenera hybrids. Lee et al. (1990) attempted this by comparing the mean performance of the tenera hybrid progenies of third and fourth generation Deli duras when crossed to the same pisiferas but the trials were planted at different periods although at similar localities. A 6.4% increase in oil yield was obtained mainly through improved oil content. Similarly, Rajanaidu et al. (1990) reported a 7% oil yield difference with tenera hybrid progenies of their first and second generation Elmina Deli duras crossed to similar pisiferas planted in the same trial. Based on the progeny-test results, selecting the best pisifera (top 15%) would give a 12% improvement (Lee and Yeow 1985; Hardon et al. 1987). Owing to the limited number (<10) of pisiferas usually available and tested, such stringent selection would seriously restrict commercial hybrid seed production. The top 30-50% selection would have been more likely in which case an improvement of around 5% would be achievable (Soh 1986). By combining the expected progress made in both the dura and pisifera parents, a gross estimate of the 10-15% breeding progress made using the modified recurrent selection scheme would not be unreasonable.
As evident, it is difficult if not futile to compare objectively the relative efficiencies of the two selection schemes. The choice depends very much on one’s bias, objectives (i.e. short or long term gains), and stage of the breeding program. We prefer a combined approach, modified recurrent selection to develop relatively unimproved populations to reduce the final size of recombinant families to go into the modified reciprocal recurrent selection program. Most mature oil palm breeding programs in the Far East are beginning to adopt the latter technique.
2. Other Breeding Methods
Backcross Breeding. This has been used in introgression work to improve a particular genotype or population. The Dumpy trait has been introgressed into the AVROS population to reduce height (Soh et al. 1981; Lee and Toh 1992). The backcross approach is also used to introgress the better oil quality traits from the Nigerian guineensis and the American oleifera materials and to restore the yielding ability and fertility of the recurrent advanced breeding parents respectively (Sterling et al. 1988; Le Guen et al. 1991; Soh et al. 1999; Sharma 2000a).
Recombinant Inbred Lines. The development of recombinant inbred lines have been proposed and attempted in oil palm (Lawrence 1983; Pooni et al. 1989). This would involve the production of dura inbred line cultivars (as the tenera is a heterozygote and the pisifera homozygote is female sterile) with oil yields superior to the best thin-shelled tenera hybrid. This is unlikely. Other disadvantages would be the possible loss of desirable genes linked to yield in the infertile gametes/zygotes in the inbreeding process, and longer time to produce a commercial variety (Soh 1987b). However, the recombinant inbred line approach combined with single seed descent may be considered for development of inbred parents for hybrid production. This will put oil palm hybrid seed production on similar footing as hybrid maize production in the generation of hybrid combinations for testing. The availability of inbred lines particularly isogenic lines will be a boon to basic studies on oil palm genetics (quantitative, molecular, physiological, biochemical) and a host of other fundamental studies and applications.
Breeding for Clonal Propagation. This would involve treating the oil palm as a clonally propagated crop such as potato, cassava, rubber, sugar cane. The breeding approach for a clonally propagated crop starts off by generating a segregating hybrid population with high mean and progeny variability. Single plant selection is followed by cycles of increasing scales of testings of the clonal offsprings, in later stages involving multi-location trials (Brown et al. 1988; Kawano et al. 1998; Tan 1987; Simmonds 1979). Currently we do not foresee this happening in oil palm as the feasibility of repeated cycles of recloning ramets (clonal offspring) is still uncertain (Soh 1998). There will be further discussions on this subject in a later section.
3. Index and BLUP Selection
In oil palm breeding, as in animal breeding, the choice of breeding parents is crucial. Firstly the number of parents that can be accommodated in the program is limited because of its perennial tree crop nature. Secondly, the hybrid production cycle is closely linked to the breeding cycle. Knowledge of the breeding values of the parents and multiple trait selection methods will help in the choice of the parents. The selection index and best linear unbiased prediction (BLUP) methods derived from application of linear statistical models which are usually associated with animal breeding (Henderson 1984) are now more commonly used in plant breeding (White and Hodge 1989) including oil palm breeding.
Selection Index. Multi-trait selection index and index selection incorporating plot and family information were found to be useful in choosing candidate palms (ortets) for cloning (Soh and Chow 1989, 1993; Soh et al. 1994a; Baudouin et al. 1994). The same approach can be used in selecting parents for breeding.
BLUP . The combining ability approach has been the usual approach adopted in selecting parents in oil palm breeding (Soh et al. 1999, Breure and Konimor 1992). Many oil palm breeding trials, however, tend to be highly unbalanced in mating as well as experimental designs due to the constraints of the crop. The combining ability approach does not readily allow objective integration and comparison of genetic information from across unbalanced experiments. In this regard, the BLUP method developed for cattle breeding, which have similar constraints, confers the following advantages (Soh 1999):
- it can handle highly unbalanced mating and experimental designs
- it can utilize information from a number of experiments even without standard check varieties
- it can utilize information from other relatives or sources
- it allows a more directed approach in breeding, breeding towards an ideal or optimum genotype and breeding towards a target environment
- it allows refinement as better genetic information becomes available
- it can handle multiple traits simultaneously
Essentially, the BLUP approach removes environment and replicate effects as “nuisance” fixed effects in order to estimate the random genetic effects e.g. breeding values or additive genotype. Soh (1994) pioneered the use of BLUP in oil palm breeding by estimating the breeding values of pisifera parents from a very unbalanced set of tenera hybrid trials and predicted increased usage of this technique. BLUP has now been used not only to estimate breeding values of parents but also in predicting hybrid performances in oil palm (Purba et al. 2000) as have been done in maize (Bernado 1994, 1995, 1996) and sugarcane (Chang and Milligan 1992a,b).
C. In Vitro Methods
1. Commercial clonal propagation. Commercial oil palm planting materials being a mixture of hybrids from non-fully inbred parents have considerable between palm genetic variability arising from between and within family genetic variabilities depending on the relatedness and inbred status among dura and pisifera parents. Individuals could vary by more than 30% of the mean yield of the hybrids (Hardon et al. 1987; Meunier et al. 1988). Although the differences might not be entirely genetic, nevertheless they provided the rationale and impetus in the 1960s to develop the in vitro propagation of the oil palm, which has no natural means of vegetative propagation. The oil palm, being a monocot and a perennial tree, was considered a recalcitrant species for in vitro propagation. However, with concerted research efforts the first in vitro oil palms were achieved by the mid 1970s (Jones 1974; Rabachault and Martin 1976; Lioret and Ollagnier 1981). Rohani et al. (2000) and Krikorian and Kann (1986) have reviewed the details and historical development of the oil palm tissue culture techniques.
However, large-scale commercial propagation and plantings of proven tenera clones have yet to take-off despite more than two decades of research and development (R&D) work involving the setting up of more than 20 tissue culture laboratories worldwide with expenditures running into tens of millions in clonal propagation for trial and pilot commercial test plantings. For successful large-scale commercial propagation of proven tenera clones, a number of critical issues need to be resolved including somaclonal variation, cloning efficiency, ortet (parent palm of clone) selection efficiency, feasibility of recloning and the liquid suspension system, clonal field evaluation requirements and expected genetic progress.
Somaclonal Variation. The first issue is the specter of somaclonal variation manifested as abnormal flower and fruit development on otherwise apparently normal palms. In the pistillate flower the vestigial androecium develops into supernumery carpels forming a “mantle” over the developing fruit (Plate IIB) similar to the rare genetic mantled or “poissoni” fruit variant found in nature (Hartley 1988). The gynoecium may be fertilized or develop parthenocarpically. The feminized (carpelloid) flowers of the abnormal staminate inflorescence do not bear pollen. Severely mantled fruit bunches tend to be parthenocarpic leading to bunch abortion and palm sterility (Corley et al. 1986; Paranjothy et al. 1990). Palms bearing slightly to moderately mantled bunches can recover to normal bunch bearing (Ho and Tan 1990; Durand-Gasselin et al 1995; Duval et al. 1997). Susceptibility to the abnormality varies between and within clones and the risk appears to increase with extended culture, the earliest batches of ramets (clonal plants) tended to be mantle free (Paranjothy et al. 1995b). The mantled fruit phenomenon was experienced by all laboratories. As some clones can be severely mantled with susceptibility apparent only at the fruiting stage (2-3 years after field planting), confidence in mass propagation was seriously eroded. Speculations were made on the physiological, epigenetic (gene expression) and genetic bases, including the involvement of cytoplasmic or organelle DNA e.g. mitochondria, of the mantle somaclonal variant (Corley et al. 1986; Soh 1987a; Jones 1995; Paranjothy et al. 1995b). Mantling was found to be apparently sexually transmissible (Rao and Donough, 1990) but this still did not rule out the epigenetic basis. Use of fast growing instead of nodular callus (Duval et al 1988; Marmey et al. 1991) and/or phytohormones such as cytokinins (Agamathu and Ho 1992; Besse et al. 1992; Jones et al. 1995, Ng et al. 1995) in the proliferation stage was suggested as the culprit but this soon proved untenable as laboratories which did not use either still produced mantled ramets.

Plate II. B) Parthenocarpic mantled fruits.
As empirical approaches to research the causal factors through protocol manipulations appear cumbersome, if not imprecise, the alternative molecular approach was initiated. Research on the molecular basis of the mantle somaclonal variation using molecular marker techniques e.g. protein, isozymes, restricted fragment length polymorphism (RFLP), random amplified polymorphic DNA (RAPD), amplified fragment length polymorphism (AFLP), differential display, has been contracted out to the laboratories of six international centers of excellence by MPOB besides in-house efforts (Cheah et al. 1999; Chowdhury 1995; Paranjothy et al. 1995a; Sharifah et al. 1999). The CIRAD (Centre de Cooperation Internationale en Recherché Agronomique pour le Development) group in Montpellier, France has also been involved in similar research (Marmey et al. 1991; Rival et al. 1997; 1998a,b; Jagligot et al. 2000; Morcillo et al. 2000). Preliminary findings implicated an epigenetic basis (also possibly a genetic basis), mediated through methylation perhaps involving retro-transposons, affecting the expression of homeotic (flowering) genes (Rival et al. 1997; Shah et al. 1999; Rajinder et al 2001). Methylation has been implicated in somaclonal variation of other crops (Phillips et al. 1994; Tsaftaris and Polidorus 2000). The development of a diagnostic tool is the ultimate objective although it looks elusive at the moment as putative molecular markers isolated so far appeared to have specific rather than general applicability on clone or culture basis. Meanwhile, an empirical approach would be to look for genotypes which are amenable to cloning and resistant to somaclonal variation and this could be subsequently followed up with the development of molecular markers or genes for these traits. Many laboratories are now reporting significantly reduced mantling rates in packages of clones planted (Table 2) and have attributed them to use of more suitable or improved protocols reinforced by stringent culture selection (Maheran and Abu Zarin 1999; Ho 2000, Soh et al., 2001). Similar experiences have been reported in banana micropropagation (Kikorian 1994).
Table 2. Mantling incidence rates in clones. Source: Soh et al. 2001.
| Year cloned |
Year |
Explant source |
No. of clones |
Clones mantledz |
No. rametsyplanted |
Mantled ramets in clones |
|
|
Range |
Mean |
||||||
|
1983/4 |
1986-91 |
Ortetx |
12 |
67 |
7,427 |
0-35.7 |
6.2 |
|
1987 |
1989-92 |
Embryos (RC)w |
17 |
12 |
6,840 |
0-72.0 |
1.1 |
|
1987/88 |
1992-95 |
Seedlings (RC) |
63 |
75 |
80,632 |
0-35.4 |
7.6 |
|
1992 |
1996 |
Ortet |
3 |
67 |
451 |
0-3.5 |
2.0 |
|
1993 |
1996-97 |
Ortet |
20 |
80 |
9,378 |
0-9.1 |
2.7 |
|
1994 |
1997-98 |
Ortet |
18 |
50 |
11,865 |
0-3.5 |
1.4 |
|
1995 |
1998 |
Ortet |
22 |
32 |
3,412 |
0-6.0 |
1.0 |
zA clone with any of its ramets expressing the mantle somaclonal variation is considered mantled.
yRamets = clonal plants or members of a clone.
xOrtet = parent tree of clone
wRepeated cross of superior family
Cloning Efficiency. The second issue is the inefficiency of the cloning process. Although callogenesis rates (100% palm-wise, 17% explant-wise), and percent embryogenic palms (80%) appear not to be process limiting, embryogenesis rates on callus cultures varied from 0.2-36%, averaging 4% per ortet (Table 3). With about 2000 leaf explants per ortet usually sampled, this translates to 12 leaf explants with embryos obtained per ortet on average with some having one or none and others having >120. More importantly is the capability of active proliferation and thus mass propagation of the embryos obtained. Generally about half are capable (Soh et al. 2001; Wooi, 1995), while the remaining will either persist in producing shoots at the detriment of embryo proliferation or fail to develop further. In the final analysis only about 18% of the ortets cultured can provide cultures amenable to mass propagation. Prospects of further improvement in efficiency appear limited although it may be possible by coaxing the cultures through more drastic protocol manipulations. Most laboratories are wary to deviate significantly from their established protocols for fear of increased risk of somaclonal variation and the additional tedious protocol field-tests encumbered.
Table 3. Oil palm cloning and recloning efficiencies, gelled-culture system.
Source: Soh et al. 2001.
|
Explant sourcez |
Basis |
Callogenesis |
Embryogenesis |
|
Ortet (cloning) |
Palm |
100 |
80 (43-100) |
|
Explant |
17 (12-27) |
4 (0.2-36) |
|
|
Ramet (recloning) |
Palm |
100 |
89 (77-100) |
|
Explant |
16 (11-21) |
12 (0.2-52) |
zOrtets = parent palms of clones; ramets = clonal plants
Ortet Selection Efficiency. Selecting superior palms or ortets for cloning is an inefficient process. In the breeding of many crops (Allard 1960; Simmonds 1979) and in clonally propagated crops e.g. potato, cassava, rubber (Brown et al. 1988; Kawano et al. 1998; Tan 1987), single plant selection especially for yield from a mixed seedling-derived population has been inefficient if not futile. Soh (1986) using data from trials of an advanced tenera hybrid (i.e. Deli × AVROS) population estimated that selecting the top 5% of the palms for cloning would give clones yielding 10-15% more than the mean of the hybrids and that clones yielding more than 30% were unlikely because of the low heritability and genetic variability. Also the better clones could only be identified after clonal testing. If the clones were to be compared to the improved hybrids, which would be available by the time the clonal test results are available, the advantage would be much reduced. Corley et al. (1999) was partial towards the higher estimate citing the higher heritability values obtained by Baudouin and Durand-Gasselin (1991) which were subsequently found, when the palms were older, to correspond to Soh’s estimates (Cochard et al. 1999). The elucidation of the low heritability for oil yield in these advanced tenera hybrid populations implied that the same experiences in other crops mentioned earlier would apply in oil palm (Soh 1998). This has been further supported by the clonal trial results of Donough and Lee (1995), Cochard et al. (1999), and Soh et al. (2001). In our first trial (Table 4), although four out of 12 clones derived mainly from highly selected ortets of Deli × AVROS lineage exceeded the Deli × AVROS hybrid control in mean oil yield (1992-2000), only two clones exceeded by more than 10% (111% and 118%). In our second trial (Table 5), which was a trial of clones derived from the embryos of a reproduced superior Deli × (Yangambi × AVROS) hybrid i.e. each clone represented a random ortet in the superior cross, the mean oil yield (1994-2000) was exceeded only substantially by three clones out of 10 (108%, 108%, 113%). Also from the second trial, for oil yield, the genetic variability between palms within the cross, estimated from the between clone variance (genetic coefficient of variation, CVg = 6%), was much smaller than the environmental variability between palms (CVe = 20%), estimated from the within clone variance. This implied that the genetic differences between palms would have been masked by the environmental effects. These results reaffirmed the inefficiency of ortet (individual palm) selection based on its phenotype and the dependence of good clonal tests to pick out the infrequent outstanding clones especially when clones tended to exhibit GxE effects (Lee and Donough 1993; Corley et al. 1995; Soh et al. 1995).
Table 4. Trial BCT4-89; yield performance of ortet clonesz. Source: Soh et al. 2001.
|
Clone
|
O/By
|
Fresh fruit bunch (FFB) yield |
Oil yieldw |
||||||||||
|
1992-1996 |
1997-2000 |
1992-2000 |
1992-1996 |
1997-2000 |
1992-2000 |
||||||||
|
(t/ha/yr) |
(% of control) |
(t/ha/yr) |
(% of control) |
(t/ha/yr) |
(% of control) |
(t/ha/yr) |
(% of control) |
(t/ha/yr) |
(% of control) |
(t/ha/yr) |
(% of control) |
||
|
1/3 |
26.1 |
21.2 |
92 |
33.3 |
106 |
26.5 |
99 |
5.5 |
87 |
8.7 |
102 |
6.9 |
95 |
|
2/7 |
26.5 |
20.6 |
89 |
30.5 |
97 |
24.9 |
93 |
5.5 |
87 |
8.1 |
95 |
6.6 |
90 |
|
3/12 |
27.8 |
23.9 |
103 |
36.0 |
115 |
29.2 |
109 |
6.6 |
105 |
10.0 |
118 |
8.1 |
111 |
|
5/15 |
25.5 |
17.9 |
77 |
26.4 |
84 |
21.6 |
81 |
4.6 |
73 |
6.7 |
79 |
5.5 |
75 |
|
5/17 |
28.0 |
18.3 |
79 |
29.2 |
93 |
23.0 |
86 |
5.1 |
81 |
8.2 |
96 |
6.4 |
88 |
|
5/14C |
29.4 |
21.0 |
91 |
30.4 |
97 |
25.1 |
94 |
6.2 |
98 |
8.9 |
105 |
7.4 |
101 |
|
6/22 |
31.6 |
22.6 |
98 |
33.3 |
106 |
27.3 |
102 |
7.1 |
113 |
10.5 |
124 |
8.6 |
118 |
|
7/26 |
28.4 |
21.8 |
94 |
32.4 |
103 |
26.4 |
99 |
6.2 |
98 |
9.2 |
108 |
7.5 |
103 |
|
9/28 |
25.5 |
18.6 |
80 |
33.7 |
107 |
25.2 |
94 |
4.7 |
75 |
8.6 |
101 |
6.4 |
88 |
|
10/39 |
27.9 |
17.4 |
75 |
27.8 |
89 |
22.0 |
82 |
4.9 |
78 |
7.8 |
92 |
6.1 |
84 |
|
12/84 |
29.4 |
19.2 |
83 |
30.9 |
98 |
24.3 |
91 |
5.6 |
89 |
9.1 |
107 |
7.1 |
97 |
|
11/283 |
27.6 |
19.6 |
85 |
23.8 |
76 |
21.4 |
80 |
5.4 |
86 |
6.6 |
78 |
5.9 |
81 |
|
DxP (AVROS)wcontrol |
27.1 |
23.1 |
100 |
31.4 |
100 |
26.8 |
100 |
6.3 |
100 |
8.5 |
100 |
7.3 |
100 |
|
Mean of clones |
27.8 |
20.2 |
88 |
30.6 |
98 |
24.7 |
92 |
5.6 |
89 |
8.5 |
100 |
6.9 |
95 |
|
LSD.05 |
2.6 |
2.4 |
–
|
3.3 |
–
|
2.2 |
–
|
1.0 |
–
|
0.9 |
–
|
0.6 |
–
|
zOrtet clones = clones derived from selected adult palms
yO/B = oil in fresh bunch by weight
xOil yield = FFB × O/B, mean for period
wDxP (AVROS) = a dura × pisifera hybrid of superior lineage
Table 5. Trial BCT9-91. Yield performance of embryo-derived clones of a reproduced superior dura × pisifera (Yangambi-AVROS) cross. Source: Soh et al. 2001.
|
Clone |
O/Bz |
Fresh fruit bunch (FFB) yield |
Oil yieldy |
||||||||||
|
1994-1996 |
1997-2000 |
1994-2000 |
1994-1996 |
1997-2000 |
1994-2000 |
||||||||
|
(t/ha/yr) |
(% of mean) |
(t/ha/yr) |
(% of mean) |
(t/ha/yr) |
(% of mean) |
(t/ha/yr) |
(% of mean) |
(t/ha/yr) |
(% of mean) |
(t/ha/yr) |
(% of mean) |
||
|
E283 |
27.9 |
18.5 |
105 |
31.0 |
110 |
24.0 |
107 |
5.2 |
108 |
8.6 |
110 |
6.7 |
108 |
|
E284 |
27.7 |
17.3 |
98 |
25.8 |
91 |
21.1 |
94 |
4.8 |
98 |
7.2 |
92 |
5.8 |
94 |
|
E308 |
27.4 |
15.5 |
88 |
27.1 |
96 |
20.7 |
92 |
4.3 |
88 |
7.4 |
95 |
5.7 |
92 |
|
E312 |
27.6 |
17.5 |
99 |
24.3 |
86 |
20.5 |
92 |
4.8 |
98 |
6.7 |
86 |
5.7 |
92 |
|
E348 |
25.4 |
18.0 |
102 |
29.5 |
105 |
23.1 |
103 |
4.6 |
94 |
7.5 |
96 |
5.9 |
95 |
|
E352 |
28.7 |
18.5 |
105 |
32.0 |
113 |
24.5 |
109 |
5.3 |
108 |
9.2 |
118 |
7.0 |
113 |
|
E353 |
29.0 |
17.9 |
101 |
26.2 |
93 |
21.6 |
96 |
5.2 |
106 |
7.6 |
97 |
6.3 |
102 |
|
E379 |
30.8 |
18.4 |
104 |
26.0 |
92 |
21.8 |
97 |
5.7 |
116 |
8.0 |
103 |
6.7 |
108 |
|
E380 |
28.0 |
16.6 |
94 |
29.3 |
104 |
22.2 |
99 |
4.6 |
94 |
8.2 |
105 |
6.2 |
100 |
|
E383 |
23.5 |
18.4 |
104 |
31.2 |
111 |
24.1 |
108 |
4.3 |
88 |
7.3 |
94 |
5.7 |
92 |
|
Mean |
27.6 |
17.7 |
100 |
28.2 |
100 |
22.4 |
100 |
4.9 |
100 |
7.8 |
100 |
6.2 |
100 |
|
LSD.05 |
1.6 |
3.0 |
–
|
3.3 |
–
|
2.7 |
–
|
1.0 |
–
|
0.9 |
–
|
0.7 |
–
|
zO/B = oil in fresh bunch by weight
yOil yield = FFB x O/B, mean for period
Alternative Approaches in Clonal Propagation. A number of analyses incorporating secondary trait, plot and family information in selection indexes had been carried out to improve ortet selection efficiency. Family information was found to be most important (Soh and Chow 1993; Baudouin et al. 1994, Soh et al. 1994a), lending support to two other alternative approaches to oil palm cultivar development using clones proposed earlier by Soh (1986, 1987a). Instead of cloning elite tenera palms which appeared inefficient if not elusive, we could clone a sample of the reproduced seedlings/palms of the best proven cross, or clone one or both parents of the best proven cross which could then be used to produce semiclonal (one clonal parent and one sexual parent) or biclonal (both clonal parents) hybrid seeds.
The rationale for the two other alternative approaches were as follows:
-
Both approaches would capture the best family oil yield performance, which would be about 10-15% above the mean of the families, as the best family would have been selected from statistically designed trials with replicated plots as compared to selecting individual palms in the original elite tenera palm cloning approach.
-
The cloning best family approach would provide a larger clone and explant base to circumvent the ortet cloning inefficiency. In the case of seed embryos and seedlings, the early and the relative ease in cloning would be advantageous.
In the clonal hybrid seed approach, risk of clonal abnormality would be minimized as fewer ramets would be needed to serve as clonal parents and could be obtained from the early cultures that are usually less susceptible to mantling. Also deleterious recessive mutations could be masked or self-selected out in the sexual seed production process.
Commercial-scale plantings from cloning a sample of the best family approach or from mass selected ortets are increasing because of improved confidence in achieving low mantling rates with the refined laboratory protocols and good yields achievable (Khaw and Ng 1997; Maheran and Othman 1999; Wong et al. 1997,1999a). Trials of biclonal and semi-clonal hybrids are giving encouraging results in terms of negligible mantling and good potential yield (Sharma 2000; Veerappan et al. 2000; C. W. Chin pers.comm. 2001) and commercial clonal hybrid seeds should be available soon.
Commercial production of proven elite clones (Plates IIC, IID), the “holy grail” of oil palm clonal propagation effort, however, has yet to be achieved. The two alternative approaches can only recapture the best family not the best individual performance. Secondly with the still inefficient cloning technique, to achieve an annual ramet production capacity of half a million, we would need to clone 100 ortets per year. The availability of sufficient elite ortets would be limiting unless a program to create a constantly available pool of elite ortets from the reproductions of the best families has been planned earlier (Soh, 1998).
 Plate II. C) A precocious high yielding clonal palm or ramet. |
 Plate II. D). Thick mesocarp, high oil bearing fruits of a clonal palm. |
Recloning. As discussed earlier, good clonal tests are required to pick out the outstanding clones. To reproduce proven clones in reasonably large numbers, the feasibility of recloning (from ramets) must be in place as resampling (from ortets) will still be constrained in ramet production by the inefficient technique and limited explant source. The feasibility of clonal reproductions from cryopreserved somatic embryos, the alternative approach, has yet to be proven (Engelman and Duval 1986; Paranjothy et al. 1986; Dumet et al. 1993, 1994). Initial experiences with recloning were discouraging because of the higher susceptibility to mantling encountered (T. Durand-Gasselin pers.comm.1999; Wong et al 1999a), although it appeared to be more amenable giving higher embryogenesis rates (Table 3). Subsequent reclonings produced lower mantling rates (Table 6). Our latest results from 24 reclones averaged 2% mantling with only two clones having relatively higher mantling rates (about 10%). A possible explanation of the decreasing mantling susceptibility with the later reclones could be the “wearing-off” of the carry-over epigenetic effects in the older ramets sampled. Recloning now appears feasible.
Table 6 . Mantling incidence rates in reclones. Source: Soh et al . 2001.
|
Year recloned |
Year planted |
No. primary clones recloned |
No. primary rametszrecloned |
Reclonesy of primary clones mantledx |
Reclones of |
No. of secondary ramets planted |
Mantled ramets in reclones |
|
|
Range |
Mean |
|||||||
|
1989 |
1993-95 |
3 |
6 |
100 |
100 |
35,330 |
2.9-14.1 |
11.3 |
|
1991 |
1994-97 |
3 |
8 |
100 |
100 |
7,853 |
2.3-9.2 |
4.9 |
|
1992 |
1995-97 |
5 |
8 |
100 |
75 |
1,016 |
0-5.2 |
3.2 |
|
1993 |
1997 |
4 |
4 |
50 |
50 |
478 |
0-3.6 |
1.2 |
|
1994 |
1997-98 |
10 |
24 |
80 |
67 |
2,779 |
0-12.7 |
2.1 |
zRamets = clonal plants
yReclones = clones obtained from cloning ramets
xClones with any of its secondary ramets (from recloning) mantled.
wRamets with any of its secondary ramets (from recloning) mantled.
Liquid Suspension Culture. With the feasibility to reclone, the prospect of mass propagation has improved. However, the conventional gelled-culture protocol involving proliferating polyembryogenic cultures with asynchronous development on which recloning has been based is still very inefficient in terms of ramet production rates and labor and space utilization (Plate IIE). It also offers limited scope for improvement and automation thus constraining the level of mass propagation. The latest development in the liquid suspension culture technique may solve this problem (De Touche et al. 1991; Duval et al. 1995; Texeira et al. 1995; Wong et al. 1999b). High efficiency has been achieved in our liquid culture system. About 80-90% of the embryogenic palm (ortet, ramet) cultures and 60-90% of their embryogenic callus cultures proliferated in the liquid system (Table 7) and most importantly, at much higher rates (6-7 times per monthly subculture) than in the gelled system (two times per bimonthly subculture). Very high success rates have also been achieved at shoot regeneration of the embryos and subsequent root induction on gelled medium (Plate IIF). Synchronized culture development is the hallmark of the liquid system and this has been largely achieved although further improvement in synchronization of embryo germination is still possible.
 Plate II. E) Polyembryogenic cultures with asynchronous development in gelled medium. |
 Plate II. F) Plant development from somatic embryos in gelled medium. |
|
Explant sourcez |
Basis |
Embryogenic callus proliferation |
Shoot conversion from embryo |
|
Ortet (cloning) |
Embryogenic palms |
83 |
94 |
|
Embryogenic callus |
63 |
95 |
|
|
Ramet (recloning) |
Embryogenic palms |
93 |
100 |
|
Embryogenic callus |
95 |
100 |
z Ortets = parent palms of clones; ramets = clonal plants
Predisposition to higher risk of somaclonal variation has been attributed to the liquid culture technique in other crops (George 1993) and also in oil palm (Y. Duval, pers.comm. 1993). The latest results from our liquid-cultured ramet plantings, for both clones and reclones, did not indicate increased risk of mantling. The few mantling incidences observed occurred mostly at much higher levels of ramet production (possible with the liquid culture method) as compared to the experiences with the gelled-culture technique (Tables 8, 9, 10, 11). CIRAD has apparently observed similar results on smaller sample test plantings (A. Rival pers. comm.. 2001). With the latest advances in the recloning and the liquid suspension culture techniques, the feasibility of commercial mass production of proven elite tenera clones is at hand.
Table 8. Mantling incidence rates in clones, liquid suspension system. Source: Soh et al. 2001.
| Source |
No. clones planted |
No. with bunches |
Normal |
Mantledz |
Mixedy |
|||
|
(No.) |
(%) |
(No.) |
(%) |
(No.) |
(%) |
|||
| Clones |
16 |
16 |
10 |
63 |
4 |
25 |
2 |
13 |
| Reclones |
16 |
16 |
11 |
69 |
3 |
19 |
2 |
13 |
| Total |
32 |
32 |
21 |
66 |
7 |
22 |
4 |
13 |
yMixed = clones/reclones having mixture of normal and mantled embryo lines
zA clone/reclone with any of its clonal plants mantled is considered mantled.
Table 9. Mantling incidence rates in embryo lines, liquid suspension system. Source: Soh et al . 2001.
| Embryo line |
No. planted |
No. with bunches |
Normal |
Mantledz |
||
|
(No.) |
(%) |
(No.) |
(%) |
|||
| Clones |
24 |
24 |
17 |
71 |
7 |
29 |
| Reclones |
23 |
23 |
17 |
74 |
6 |
26 |
| Total |
47 |
47 |
34 |
72 |
13 |
28 |
zA clone/reclone with any of its clonal plants mantled is considered mantled
Table 10. Mantling incidence rates of ramets, liquid suspension system. Source: Soh et al. 2001.
| Rametz type |
Ramets planted |
No. ramets with bunches |
Mantling incidence |
|
|
(No.) |
(%) |
|||
| Primaryy |
1536 |
1249 |
36 |
2.9 |
| Secondaryw |
1758 |
1394 |
57 |
4.1 |
| Total |
3294 |
2643 |
93 |
3.5 |
zRamets = clonal plants
yPrimary ramets = ramets from cloning
xSecondary ramets = ramets from recloning
Table 11. Ramet production and mantling rates of clones in gelled vs. liquid culture systems at various subculture levels. Source: Soh et al. 2001.
|
Clone no. |
Gelled-culture system |
Liquid suspension system |
||||
|
Subculture |
No. shoots obtained |
Mantled ramets |
Subculture no. |
No. shoots obtainabley(million) |
Mantled ramets |
|
|
41 |
11 |
1120 |
73 |
11 |
3 |
93 |
|
178-46 |
7,11,13 |
375 |
7 |
7,8,9 |
0.09 |
0 |
|
100-24 |
13, 14 |
314 |
4 |
3, 15, 16 |
0.003 |
0 |
|
154-44 |
9 |
290 |
0 |
9, 13 |
0.02 |
9 |
|
94-195 |
7 |
147 |
0 |
9 |
20 |
2 |
|
124-24 |
6 |
636 |
0 |
1,3 |
0.001 |
0 |
|
180-2 |
6,8,9 |
169 |
0 |
6, 7 |
0.01 |
0 |
|
79-24A |
14, 15 |
201 |
0 |
13, 17 |
0.4 |
0 |
|
121-29A |
6, 9 |
342 |
0 |
3, 8 |
6 |
0 |
|
176-86 |
9, 14 |
1108 |
0 |
4,8,9,14 |
1000 |
0 |
zAbout 40 ramets (clonal plants) were sample-tested per subculture.
yExtrapolated figures based on proliferation and germination rates of cultures
Field Evaluation. The oil palm industry is about to enter a new era of large-scale plantings of more genetically uniform materials in the form of clonal hybrids and clones. Nevertheless, there are still issues that need to be addressed and investigated now to ensure successful exploitation of these technological developments and minimize risk of catastrophic setbacks.
On the propagation side there is need to demonstrate that clonal hybrids do not perform poorer than sexual hybrids because of reduction in fitness of the clonal parents (Bregitzer and Poulson 1995; Phillips et al. 1994). This should be repeated with hybrids of recloned parents as there may be need to perpetuate the parents of outstanding hybrids. There is also need to know whether mantled ramet parent or normal ramet parent of mantled clones would transmit the abnormality to their hybrid progenies.
It would be useful to demonstrate the heritability of clonability and susceptibility/resistance to clonal abnormality and their repeatability in clones and reclones. It would also be essential to identify the predisposing environmental factors. Solving these issues, perhaps with the development and assistance of molecular markers, would facilitate the selection (perhaps also breeding) of amenable and resistant genotypes for cloning. We also need to establish, if feasible, safety limits for mantling risk and reduced fitness for cycles of recloning and for levels of production especially for the liquid culture technique. Molecular markers would have an important role here. Prospects for optimization and automation of the liquid culture protocol are still good and the development of synthetic seed might be more efficient than dealing with regenerated plantlets.
On the field testing and planting side, clonal hybrids and clones being genetically more homogeneous and discrete than mixed hybrids, would respond differently to different environmental factors e.g. location, planting density, fertilizer requirement, and abiotic (soil, water, mineral) and biotic (pest, disease) stresses i.e. exhibit GxE effects (Corley and Donough 1992; Donough et al. 1996; Smith et al. 1996). Adaptability trials are thus mandatory. For clonal hybrids, the choice of the hybrids should be based on GxE tests of the original sexual parental hybrids. In the case of clones which more prone to GxE effects and where ortet selection is inefficient, repeat GxE tests may be necessary to pick out the superior clones. In both cases to reduce the time and effort in GxE testing for a perennial tree crop, the selection of a minimum sample of environments that can discriminate the differential responses of the genotypes yet representative of the major environments where the genotypes are to be grown needs urgent consideration and investigation. This would be especially so for clones to reduce the need for repeat cycle tests and the number of candidate clones in each subsequent cycle. Use of molecular marker and quantitative trait loci (QTL) assisted selection has often been suggested but with a quantitative trait susceptible to GxE effects such as yield, the task would be difficult if not daunting.
As is common for successful cultivar development in most crops, there is always the need to produce or reproduce superior families preferably of different genetic origins and in larger numbers to provide the ortets for cloning to feed the subsequent clonal testing and selection program. With successful cloning, large plantings of genetically homogeneous materials of restricted genotypes can be foreseen and this would lead to genetic vulnerability of the crop to epiphytotics, pest outbreaks, inadequate pollination and moisture and mineral deficiencies. Thus, research is required for the packaging of clones and their field planting arrangements to reduce the genetic vulnerability and to also synchronize the cropping pattern with the future mechanized harvesting, and new crop management systems.
Genetic Progress. There have been reports of markedly superior yields of clones over tenera hybrids. Comparisons have sometimes been made against unspecified commercial hybrid materials with the attending errors due to between and within material variabilities and sampling. Even when individual crosses were used, the relative performances of the standard crosses within the respective proprietary commercial hybrid materials were not known. The comparisons of Cochard et al. (1999) were more objectively based having link crosses representing the first generation commercial hybrid material from their reciprocal recurrent selection program. The relative performance of the second generation hybrids with respect to the first generation hybrids was also known. The advantage of cloning improvement over breeding improvement should be based on the comparison between those of the clones and the second generation hybrids as discussed earlier. Soh (1998) suggested use of the reproduced hybrid family of the ortet as the control since the relative performance of the family in the hybrid progeny test, which represents the commercial hybrids, is usually known. This will also enable studies on selection response and selection efficiency to be made.
In terms of the relative genetic progress achievable by breeding and cloning, Jacquemard and Durand-Gasselin (1999) basing on their reciprocal recurrent selection scheme and their higher heritability estimate projected a 10-15% oil yield advantage of clones over seeds (conventional, clonal) for 7-8 years. The projection by Soh et al. (2000) was more conservative at 5-10% advantage for about 5 years with their modified recurrent selection based program (Fig. 4).
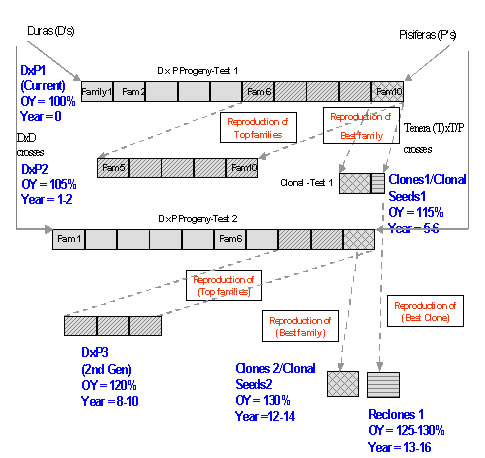
Fig. 4. Expected Breeding & Cloning Progress For Oil Yield (OY) .
DxP1 = current commercial tenera hybrids
DxP2 = reproduction of top DxP1 hybrids
DxP3 = reproduction of top families of improved second breeding cycle hybrids
Clones1, 2 = clones from selected palms of the best families of the first and second cycle hybrids
Clonal Seeds1,2 = reproductions of the best families of the first and second cycle hybrids using clonal parents
Reclones1 = reproduction of the best Clones1 clones
Assumptions (discussed in text):
Breeding progress = 15% per generation of 8-10 years;
Progress from reproduction of top families = additional 5%, obtainable 1-2 years after progeny-test results;
Progress from reproduction of best families by clones/clonal seeds = additional 15%, obtainable 5-6 years after clonal test results;
Progress reproduction of best clones i.e. reclones = additional 10-15%, obtainable 3 years after clonal test results.
Selection and reproduction of the top tenera hybrids i.e. DxP2 would give about 5% oil yield increase from the current hybrids (DxP1) within one to two years of obtaining the DxP Progeny Test1 results. Reproduction of the top hybrids (DxP3) from DxP Progeny Test2 results of the next breeding cycle hybrids eight to ten years later would give another 15% progress, 10% from improved second generation hybrids and 5% more from the top hybrids. Reproduction of the best hybrid family by cloning a sample of the family or from clonal seeds (hybrids from clonal parents) i.e Clones1 / Clonal Seeds1 available in five to six years’ time would give 15% progress. Similarly the next generation of clones (Clones2) and clonal seeds (Clonal Seeds2) available by year 12-14 would give an additional 15% progress. Reproduction of the best clone (Reclones1) in year 13-16 from the Clonal Test1 results would give an additional 10-15% progress from Clones1. Thus on average clones would have an advantage of 5-10% over improved hybrids for about five years.
These were at best rough projections as sufficient data on GxE effects and somaclonal effects on production traits in clones and reclones (Phillips et al. 1994; Bregitzer and Poulson 1995), although reported in other crops, are as yet unavailable in oil palm. Genetic progress also depends on the relative selection pressures placed on the hybrid families and clones for commercial reproduction and the genetic variabilities of the base hybrid populations.
2. Clones For Basic Studies And Other Applications
Besides commercial propagation, clones are useful for basic studies in genetics and physiology and other applications. Clones which allow replication of individual plant genotypes have useful application in basic studies on quantitative genetics e.g. partitioning between genetic and environmental variability between palms, estimation of heritability, and comparison of selection responses to different selection methods (Soh 1999, 2000). Clones will also be useful for physiological studies on crop yield potential (Corley and Donough 1992; Smith et al. 1996), the physiological bases of GxE responses and for plant developmental studies both at the plant and the in vitro level particularly with the more homogeneous suspension cultures.
Genetic transformation depends on a reliable clonal propagation technique (Parveez et al. 1998). Protoplast cultures were attempted to facilitate genetic transformation but plant regeneration has yet to be achieved (Bass and Hughes 1984; Budiani et al. 1998; Sambanthamurthi et al. 1996). Parveez et al. (1998) successfully used the biolistics approach with polyembryogenic cultures. Anther/microspore culture is useful in initiating haploids for dihaploid hybrid breeding but dihaploids have yet to be obtained in oil palm (Tirtoboma 1998).
D. Molecular Breeding
1. Marker-Assisted Selection. Marker-assisted selection has long been a quest of plant breeders (Allard 1960). With the development of molecular techniques, molecular markers closely linked to quantitative trail loci (QTL) controlling desirable traits are being sought. Such markers enable earlier and more precise selection, which is particularly advantageous for perennial tree crop breeding. The development of genetic linkage maps and markers associated with the desirable traits is a prerequisite. A program to map the oil palm genome using RFLP, AFLP, and microsatellite probes has been initiated (Mayes et al. 1997a; Cheah et al. 1999; Cheah 2000). Mapping the shell gene using bulk segregant analysis is the first obvious objective (Mayes et al. 1997b; Moretzsohn et al. 2000). Rajinder and Cheah (1999) using the bulk segregant and pseudo-test cross analyses on the selfed progenies of a high iodine value Nigerian breeding parent and a tenera clone and the progenies of a oleifera × quineensis interspecific hybrid were able to locate the locus for the virescens fruit and also found putative QTL for carotene content and the clonal mantling abnormality. The generation of high density maps for positional cloning and efforts at rapid gene discovery using expressed sequence tags (EST) are underway. Use of the GISH technique to discriminate the guineensis and oleifera chromosomes in oleifera × guineensis hybrid backcross breeding has been mentioned earlier. Other traits of interest for marker-assisted selection would include dwarfness, disease resistance and amenability to tissue culture.
While molecular marker assisted selection, which is essentially genotypic selection, is an exciting development for breeding, its success to date has been below expectation even in annual crops. Marker-assisted selection has appeared to be effective in early segregating generations, a combination of marker-assisted selection and phenotypic selection better for subsequent generations, and only phenotypic selection effective in later cycles (Stuber et al. 1999). Likewise, Bernado (2000), in a simulation study to evaluate the impact of genomic knowledge (sequence and function) of the genes controlling a quantitative trait, found that the incorporation of gene information with phenotypic selection using BLUP was most useful in selection of traits with few loci but not for traits with many loci e.g yield. Lack of well saturated uniformly spaced markers, QTL with small effects generally well distributed throughout the genome, which may be susceptible to GxE effects, level of linkage disequilibria, sample sizes, breakdown of favorable epistatic complexes, and multicollinearity (lack of independence among the factors whose effects are being measured), are some the factors considered responsible for the limited success of marker-assisted selection (Stuber et al. 1999; Bernado 2000). Undeniably, marker-assisted selection will be most useful in backcrossing and introgression programs. Molecular markers and probes have also been used to discriminate oil palm breeding populations, newly collected semi-wild populations (Shah et al. 1994; Mayes et al. 1997b; Kulratne et al. 2000; Purba et al 2000b; Rajanaidu et al. 2000), and clones (Cheah and Wooi 1995). They have also been used to determine the coancestry of parents for quantitative genetic analyses (Purba et al. 2000a).
2. Transgene Technology
Transgenic oil palms containing the Basta® resistance gene are available at the nursery seedling stage. The transgenes for high oleic acid production, which is the objective, have yet to be inserted (Parveez et al. 1998). High oleic acid was selected as the transgene of choice because a high oleic palm oil would enable it to compete with the liquid cooking oil market in temperate countries. Furthermore, oleic acid is a good industrial chemical feedstock. The approach (Cheah 2000; Shah et al. 2000) adopted involves down regulating the palmitoyl-ACP thioesterase, by using an antisense gene and up regulating the ß-ketoacyl ACP synthase II (KASII) activities, by using a strong promoter, to maximize oleic acid production at the expense of palmitic acid (Fig. 5) The thioesterase, KASII, and desaturase genes and the promoters for expression of the genes in the mesocarp tissues have been obtained and transformation will follow.
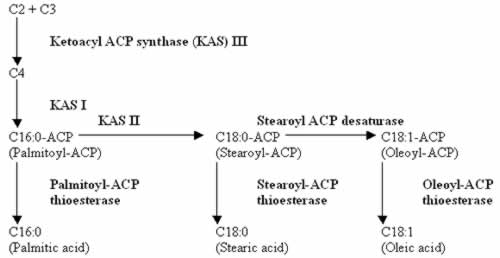
Figure 5 . Pathway to achieving high oleic oil. Palmitoyl-ACP thioesterase is down-regulated using an antisense gene and KAS II is up-regulated using a strong promoter to maximize oleic acid at the expense of palmitic acid. Source: Cheah, 2000
Plant Breeding: Germplasm Resources
A. Genetic Base and Diversity
Expeditions and collections made by early oil palm researchers at the various centers in West Africa e.g Nigeria (Calabar, Ufuma), Ivory Coast (La Me), Zaire (Sibiti, Yangambi,), especially after World War II formed the genetic base and diversity of the respective oil palm breeding programs (Rosenquist 1986; Hartley 1988). There were subsequent genetic material exchanges for breeding purposes. Most of the oil palm breeding populations were descended from one or a few palms and were termed by Rosenquist (1986) as breeding populations of restricted origins. A brief description of some of the major breeding populations is given below:
1. Deli. This is the thick-shelled dura variety derived from the original four Bogor palms in Java. Distribution of the subsequent progenies to other countries followed by local selection led to the development of the Elmina, Serdang Avenue, and Ulu Remis Deli dura subpopulations/selections in Malaysia and the Dabou and La Me dura subpopulations/selections in Ivory Coast. The Ulu Remis Deli duras were most widely distributed. The rather uniform high yielding Deli population led to the speculation of a common progenitor for the four Bogor palms. Deli duras provide the mother palms for almost all major oil palm commercial hybrid seed production programs. The Dumpy and Gunung Melayu palms are short variants of the Deli;
2. AVROS . Seeds from the Djongo (best) palm at Eala Botanical Garden in Zaire were obtained and planted in 1923 by Algemeene Vereniging van Rubberplanters ter Oostkust van Sumatra (AVROS) at Sungai Pancur to give rise to the well-known SP540 tenera palm. Crossing with the teneras at Bangun Bandar Experimental Station and subsequent backcrossing to the SP540 selfs resulted in the AVROS population. AVROS pisiferas are noted for their vigorous growth, precocious bearing, thin shell, thick mesocarp and high oil yield conferring attributes. Major commercial hybrid seed production programs in Colombia, Costa Rica, Indonesia, Malaysia, and Papua New Guinea are based on Deli dura x AVROS pisifera lineage;
3. Yangambi . The Institut National pour l’Etude Agronomique du Congo (INEAC) started the breeding program at Yanagambi, Zaire with open-pollinated seeds from the Djongo palm and from teneras in Yawenda, N’gazi and Isangi. Their breeding led to the development of the Yangambi population characterized by its excessive vigor, bigger fruit and high oil yield conferring attributes. The Yangambi population is featured in many breeding and seed production programs worldwide;
4. La Me . The Institut de Recherches pour les Huiles et Oleagineux (IRHO) developed the La Me population from 21 tenera palms, particularly the Bret 10 palm, grown from seeds collected from the wild groves in Ivory Coast. Pisiferas and teneras derived from the L2T tenera palm are used in breeding and seed production programs in West Africa and Indonesia. La Me teneras and their progenies are characteristically smaller palms with smaller bunches and fruits but they appear to be more tolerant of sub-optimal growing conditions.
5. Binga . This subpopulation is derived essentially from the F 2 and F 3 Yangambi progenies planted in Binga Plantation, Yangambi, Zaire. Palms Ybi 69MAB and Bg 312/3 are the parent palms of breeding interest.
6. Ekona . The Ekona population is derived from the wild palms in the Ekona area of Cameroon and bred further in the Unilever plantations of Cowan Estate, Ndian Estate, and Lobe Estate. Progenies from palm CAM. 2/2311, noted for its high bunch yield, good oil content and wilt resistance, have been distributed to Costa Rica and Malaysia;
7. Calabar . The breeding populations of the Nigerian Institute for Oil Palm Research (NIFOR) are much broader based with collections from Aba, Calabar, Ufuma, Umuabi. Progenies from the Calabar selections were most interesting and have been distributed to Costa Rica, Ghana, Indonesia, Ivory Coast and Malaysia. Palm NF 32.3005 was featured in most of the distributions.
Despite the reasonably diverse original genetic base, the bulk of the current commercial oil palm hybrid seed production and the breeding programs that support them are still based on the Deli and the Yangambi/AVROS and La Me breeding populations which can be traced to one or two palms from the natural groves. Because of the high selection intensities practiced and the need to retain the proven pedigrees of the hybrids for commercial reasons, genetic variability has been rapidly diminished as stressed by a number of researchers (Thomas et al. 1969; Ooi et al. 1973; Hardon et al. 1987). Prompted by this and the increasing importance of the crop, Malaysia with the cooperation of Nigeria under the auspices of the International Board For Plant Genetic Resources made the first systematic large-scale collections in the semi-wild groves in the oil palm belt of Nigeria (Rajanaidu et al. 1979; Obasola et al. 1983) with the objectives of genetic conservation and base broadening. The study on the genetic structure of the natural populations from the Nigerian collection revealed greater variability within families and between families than between populations. This guided the approach made in subsequent collections in Angola, Cameroun, Gambia, Guinea, Madagascar, Sierra Leone, Tanzania, and Zaire (Rajanaidu and Rao 1988). There was also a limited attempt to capture different ecotypes. The collected materials were planted and maintained as living collections in Malaysia with a sample retained by the host country for each collection (Rajanaidu 1990b; Rajanaidu and Jalani 1994) although long term conservation through cryopreservation and in vitro methods has also been attempted (Engelmann and Duval 1986; Paranjothy et al. 1986; Rohani and Paranjothy 1995).
Collections have also been made for E. oleifera in South America (Brazil, Colombia, Costa Rica, Honduras, Panama, and Surinam as well as for other oil bearing palms e.g. Bactris gasipaes (Pejibaye), Jessenia-Oenocarpus,, Orbignya martiana (Babassu), which have unusual fatty acid composition and other uses. These could be used in future genetic modification of palm oil.
B. Base Broadening
Private breeding programs although obliged to broaden the genetic base of their breeding populations to ensure breeding progress are generally reluctant to outcross with less advanced breeding materials and thus tend to end up with related materials. High unsaturated fatty acid content, large kernel, and dwarf high yielding lines have been developed from the Nigerian collection and distributed to the private industry breeders. Private breeders are reluctant to exploit them directly in commercial hybrid seed production although they have been urged to do so (Rajanaidu et al. 1999, 2000), but prefer to introgress the desirable features into their advanced breeding parents to maintain the integrity, uniformity, and consistency of their commercial hybrids (Sharma 1999; Soh et al 1999). These qualities may not be assured in the commercial hybrids developed directly from the early selections of the semi-wild Nigerian collections.
C. Genetics and Cytogenetics
Most oil palm breeding programs being commercially oriented cannot afford to undertake basic genetic studies. Thus such studies tend to be incidental, derived from breeding trials and seldom meet the basic assumptions required for genetic analyses.
1. Qualitative Traits . The revelation of the simple Mendelian inheritance of the shell thickness trait in the palm fruit in the 1940s by Bernaert and Vanderweyen (1941) provided the basis of modern oil palm breeding and hybrid seed production schemes (Plate 1A, Figs. 2, 3). The dura palm is homozygous dominant for the thick shell trait while the pisifera palm is the shell-less recessive homozygote which is usually female sterile as its pistillate inflorescence tends to abort. The dura × pisifera hybrid is the thin-shelled tenera heterozygote and is the commercial variety. The duras which are the female parents in hybrid seed production are usually regenerated (100%) from dura × dura crosses while the pisifera male parents are regenerated from tenera × tenera (25% pisifera segregants) and tenera × pisifera (50% pisifera segregants) crosses. Dura × tenera crosses used in progeny-tests segregate 1:1 for dura and tenera palms. Fertile pisiferas do occur occasionally and fertile pisifera × fertile pisifera crosses produce 100% pisifera progenies. Although the presence of shell is under monogenic control, the thickness of the shell was purported to be modified by minor genes (Van der Vossen 1974). Okwuagwu (1988) and Okwuagwu and Okolo (1992,1994) postulated maternal inheritance of kernel size with the involvement of a kernel-inhibiting factor.
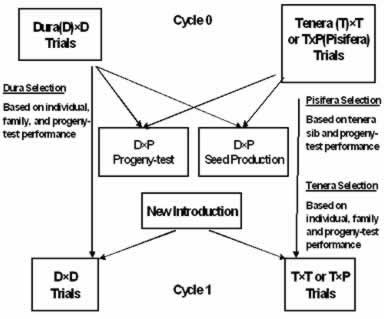
Fig. 2. Modified recurrent selection scheme.

Fig. 3. Modified reciprocal recurrent selection scheme. To sustain genetic variability for longer term improvement a recombinant phase involving outbred parents is run in parallel in a similar manner.
A number of other qualitative traits have also been reported from observations and genetic analyses to be simply inherited (Hartley 1988) e.g. crown disease, leaf form (idolatrica = fused pinnae), fruit shape (mantled), and fruit color (nigrescens = black unripe, virescens = green unripe, albescence = whitish unripe). Although the Dumpy trait appeared to be simply inherited, observations on segregating progenies did not confirm it (Soh et al. 1981). The mode of inheritance for tolerance to fusarium wilt has yet to be elucidated (De Franqueville and Renard 1990).
2. Quantitative Traits . Estimations of genetic variance and heritability have been attempted by oil palm breeders to provide information to guide them to increased breeding efficiency. This involves information on the existence of sufficient genetic variability, the choice of the population to improve and the adoption of appropriate breeding procedure to obtain the best selection response (Soh and Tan 1983). North Carolina Model 1 (hierarchical) and Model 2 (factorial) mating designs and the parent-offspring method are commonly used to estimate genetic variance and heritability in oil palm. Heritability estimates for various agronomic traits have been compiled by Soh and Tan (1983) and Hardon et al. (1985). With the use of data from breeding experiments derived from selected parents from very restricted populations, some of the assumptions such as random choice of parents, no correlation of genotypes at separate loci, and a definite reference population, necessary for valid quantitative genetic analyses could not be met and the biases incurred could not be estimated. These estimates merely serve as a guide for selection decisions as any estimate of heritability is unique i.e. dependent on the population and environment sampled and the estimation method used. For selection decisions or for prediction of selection response for a particular population and environment, specific heritability estimate obtained from the population concerned should be used. In lieu of reliable specific heritability estimates or if the responses are for different environments, average heritabilities may be substituted.
3. Cytogenetics . Cytological analyses by Maria et al . (1995, 1998a,b) on the karyotypes of E. guineensis and E. oleifera confirmed that both species have 2 n =32 chromosomes and had similar chromosome arm lengths for the three chromosome groups. These factors might explain the crossability of the two species and the partial fertility of their hybrids (Hardon, 1986). Genomic in situ hybridization (GISH), in which total genomic DNA is used as a probe for hybridizing to chromosome spreads, has been used to separate the parental chromosomes in oleifera × guineensis hybrids and would be useful in backcross breeding programs. There are two estimates of the oil palm genome size: Jones et al. (1982) estimated 1900 megabase pairs using Fuelgen microdensitometry while Rival et al. (1997) estimated 3400 using flow cytometry, the discrepancy attributed to the different methods used.
Oil Palm: Plant Breeding
Oil Palm Genetic Improvement
Introduction
Germplasm Resources
Improvement Objectives
Breeding Techniques
Future Prospects
A. Origins
The African oil palm (Elaeis guineensis Jacq.), which is the oil palm of commerce, is endemic to tropical Africa stretching from the Atlantic coast (Guinea) in the west to Zanzibar and the island of Madagascar in the east and from the sub-Sahara in the north (Senegal, 16 °N) to the south of Angola (15 °S) bordering Namibia (Hartley 1988; Latiff 2000). The center of origin and diversity appears to be concentrated in the tropical rain forests of west and central Africa (Angola, Cameroon, Congo, Ghana, Ivory Coast, Nigeria and Zaire). They occur naturally as semi-wild groves in swamps and riverine forest fringe areas in the plains usually close to settlements but are also found on drier and higher grounds up to the altitude of 1500 m above sea level. The American oil palm (Elaeis oleifera) which also produces an oil in smaller quantities consumed by the natives and is of interest in breeding because of some of its desirable attributes, however, is endemic to tropical Latin America stretching from Mexico in the north to the Amazonas of Bolivia, Brazil, Colombia and Peru in the south and straddling the Pacific and Atlantic coasts. They also occur naturally in groves in open grasslands and riverine areas and their distribution appears to be associated with indigenous Indian migratory movements (Santos et al. 1986). The distribution of the groves tended to be more discontinuous.
The genus Elaeis, which belongs to the Cocoideae subfamily of the Palmaceae, has an American or African center of origin. The two oil palm species were presumed to have diverged when the American and African continents drifted apart in prehistoric times (Zeven 1965). There are two lesser-known species of Elaeis: E. odora (South America) and E. madascariensis (Africa).
B. Crop status
Crude palm oil extracted traditionally from the fruit pulp of the semi-wild oil palms was the source of dietary fat and certain vitamins (A and E) of the indigenous populations of tropical Africa and America. Interest in the oil palm as a crop arose as a substitute for animal fat in the manufacture of soap, candle wax and margarine. Plantations were started in the East Indies (Indonesia, Malaysia) by the European colonists to ensure a steady supply of the raw material (Hartley 1988). Two seedlings each from the botanic gardens of Amsterdam and Mauritius, the thick-shelled or dura form (Plate 1A) derived presumably from the same palm in West Africa, were taken and grown in the Bogor Botanic Gardens, Java, Indonesia in 1848. With the expanding interest in the crop, subsequent hybridizations and selections were made from these four Bogor palms and the progenies distributed to the Deli province in Sumatra and thence Malaysia (Malaya then) to become known as the Deli dura population (Rosenquist 1986). This uniform high oil yielding Deli dura was the commercial planting material for the rapidly expanding plantations in the Far East from 1911 till the early 1960s. With the revelation of monogenic inheritance of the shell gene by Beirnaert and Vanderweyen (1941) and that the hybrid progenies of the thick-shelled dura and the shell-less female sterile pisifera are 100% thin-shelled teneras having thick-mesocarp and thence higher oil yield (Plate 1A), the change to the tenera hybrid planting material was very rapid as teneras from West Africa had been imported and bred earlier by researchers in the Far East and they became the source of the pisiferas (Hartley 1988).

Plate I. A) Thick shelled dura (D), shell-less pisifera (P), and thin shelled hybrid tenera (T) fruits of oil palm
The oil palm is the second most important oil crop next to soybean and is poised to become the dominant oil crop early in the new decade (Mielke 2000; Yusof and Mohd Arif 2000). Palm oil constitutes 19% of the world’s oils and fats production and is the dominant oil of international trade. The oil is produced from about 6 million hectares of plantations in the countries of the humid tropics i.e. Colombia, Indonesia, Ivory Coast, Malaysia, Nigeria, Thailand, with Malaysia accounting for 48% and Indonesia 31% of production. With the rapid expansion of the world population particularly in the third world countries where dietary fat intake is still very low, per capita oil and fat consumption is likely to increase tremendously and the oil palm being the most productive and profitable oil crop will continue to expand in its cultivation to meet this demand.
C. Uses
The oil palm fruit bunch produces two types of oil, “palm oil” from the mesocarp (20%) and “palm kernel oil” from the kernel (3%). Crude palm oil extracted from the sterilized mesocarp is refined, bleached from its original orange-red color, and deodorized to give refined palm oil used solely or blended with other oils in cooking oil, salad oil, margarines and spreads. The refined oil is fractionated to give olein and stearin and with further fractionation gives fatty acids and alcohols, intermediate commodities traded and used in food and oleochemical industries. Palm oil can be used as a biofuel as with other vegetable oils. About 80% of palm oil, however, is used in the food industries although its other uses are increasing. Palm kernel oil is a competitor for coconut oil and has more uses in the oleochemical industries. Fig.1 is a summary chart of the fractionated products and their uses.

Enlarge Fig. 1: Uses of palm oil
Secondary and waste products from the palm oil industry are assuming economic importance prompted largely by health and environmental concerns. Carotene from the crude palm oil can be processed into Vitamin A supplement and a natural dye for snack foods e.g. instant noodles. Likewise, tocopherols and tocotrienols can be extracted from palm oil for industrial Vitamin E production (Choo 2000; Jalani et al. 1997). Kernel cake and sludge cake, wastes from the palm and kernel oil extraction processes, respectively have use as animal feed while the fruit bunch stalk fiber waste can be used directly in the plantation as an organic mulch or processed into an organic compost.
Reference
Soh, A.C., Wong, G., Hor, T.Y., Tan, C.C. and Chew, P.S. 2003. Oil palm genetic improvement. In: Janick, J. (ed.). Plant Breeding Reviews Vol. 22. John Wiley & Sons, New Jersey, pp. 165-219
Note: The full list of references quoted in this article is available from the above paper.